
Meiofauna community structure and a new recorded species of free-living marine nematodes in Qigu Lagoon of Taiwan
-
摘要:
本研究于2022年8月对台湾岛七股潟湖小型底栖动物的类群组成、丰度及海洋线虫种类组成等进行分析。共鉴定出5个以上的小型底栖动物类群,分别为自由生活海洋线虫、寡毛类、桡足类、涡虫类、多毛类和其他未鉴定类群,海洋线虫为数量最优势类群,占小型底栖动物总丰度的78.52%;小型底栖动物的丰度介于(4.79±2.50)~(492.04±416.79)个/10 cm2之间;本次样品共鉴定海洋线虫491条,隶属于11科21属,海洋线虫优势属为Neochromadora、Haliplectus和Terschellingia。列出游咽线虫属(Eleutherolaimus)中有效种的形态学数据,发现国内新纪录种—拟施耐德游咽线虫(Eleutherolaimus paraschneideri),并对其进行了测量,比较了其与模式种的主要差异。
Abstract:In this study, the group composition, abundance of meiofauna and species composition of marine nematodes were analyzed using investigation data of Qigu Lagoon of Taiwan in August 2022. More than five meiofauna groups and total of 491 species belonging to 11 families and 21 genera were identified, including free-living marine Nematode, Oligochaeta, Copepoda, Turbellaria, Polychaeta and other groups. Marine nematodes was the most dominant group, accounting for 78.52% of the total abundance. The abundance ranged from (4.79±2.50) to (492.04±416.79) ind./10 cm2. Dominant genera of marine nematodes were Neochromadora, Haliplectus, and Terschellingia. The morphological data of the valid species in Eleutherolaimus were collected. A new record of free-living marine nematodes species, Eleutherolaimus paraschneideri, was described. The main differences between the new record species and model species were compared.
-
Keywords:
- meiofauna /
- marine nematode /
- Eleutherolaimus paraschneideri
-
小型底栖动物是海底碎屑食物链中的重要连接者,是鱼虾幼体的食物来源,对微生物的生产起着调节作用[1]。其中自由生活海洋线虫(以下简称海洋线虫)是海洋沉积环境中数量最为丰富,分布较为广泛,物种多样性较高的一类小型底栖无脊椎动物,在底栖生态系统的能量流动和物质循环中发挥着重要的作用[1-2]。前人曾对台湾海峡及其附近海域进行过小型底栖动物的研究[3-7]。对台湾本岛小型底栖动物的研究仅见Cai等对台湾淡水河口潮间带不同生境小型底栖动物的报道[8];Xiao等描述了台湾岛西部沿海湿地拟齿线虫属(Parodontophora)、轴线虫属(Axonolaimus)和三齿线虫属(Trissonchulus)的4个新种和1个新纪录种[9-10]。
沿海潟湖是相对较浅的水域,潟湖的形成是河流从上游带着泥沙输送至河口,泥沙淤积于河口再受到浪流作用形成离岸沙洲,与陆地间围成潟湖,部分或全部与海洋封闭[11]。七股潟湖位于台湾岛西南台南市海岸,属于半封闭的沙洲型潟湖,是台湾岛最大的潟湖生态水域,也是重要的牡蛎养殖场,其离岸沙洲在台风暴潮期间肩负着防护后方水域和陆地的重大责任[12]。邱英哲等曾对七股潟湖多毛类群与环境因子之间的关系进行研究[13],但对于七股潟湖沉积物中海洋线虫的研究目前尚未见报道。本研究对七股潟湖小型底栖动物进行研究,重点对小型底栖动物的类群组成及丰度、海洋线虫种类组成进行研究,描述了游咽线虫属(Eleutherolaimus)国内1个新纪录种,以丰富该区域海洋线虫物种多样性,为评价七股潟湖沉积环境、促进海洋生物资源的可持续利用提供参考依据。
1. 材料与方法
1.1 采样方法及站位设置
2022年8月,在台湾岛西南台南市七股潟湖设置5个采样站位,分别为A、B、C、D、E点,其中A点为大寮外排靠近七股潟湖北潮口,B点靠近七股盐田海堤,C点位于红树林区,D点靠近溪河注入口,E点靠近南潮口。每个站位设置3个重复样,选择未受扰动的地点进行沉积物样品采集,用内径2.9 cm注射器改制的采样管,从表层采集深度5 cm的沉积物样品,2份样品用5%的福尔马林溶液固定,1份样品用戴斯-马丁氧化剂(Dess-Martin periodinane solution,DESS)溶液固定,常温保存带回实验室进行分析。采样站位信息及其生境见表1。
表 1 七股潟湖采样站位信息及其生境Table 1. Sampling station information and habitat of Qigu Lagoon采样站位 经度 纬度 生境 A 120.0746 °E23.1725 °N沙洲 B 120.0772 °E23.1522 °N泥滩 C 120.0871 °E23.1307 °N红树林 D 120.0881 °E23.1174 °N泥滩 E 120.0397 °E23.1057 °N沙洲 1.2 样品处理与分析
用套筛分离法进行初处理,将样品倒在由500 μm和42 μm两层网筛组成的套筛内,用经38 μm网纱过滤的自来水缓缓冲洗,去除泥沙、黏土、甲醛固定液和其他杂质等,透过套筛的水流澄清。将截留下的样品,用密度为1.15 g/mL的Ludox-TM硅胶溶液转入离心管,以
4000 r/min转速,离心10 min,重复离心两次;将上清液用42 μm孔径网筛过滤,随后将小型底栖动物装入样品瓶中,分别加入1~2滴虎红染液(0.1 g虎红染料溶于100 mL蒸馏水),摇匀后静置24 h待用。将所截留的全部小型底栖动物转移到划有等宽平行线的培养皿中,在解剖镜下(型号:Nikon SMZ 1270)对所有的小型底栖动物进行鉴定和计数。同时利用解剖针将海洋线虫全部挑出到含有95%酒精、甘油、水的混合液(V酒精∶V甘油∶V水=1∶1∶18)的凹型玻璃容器。放在干燥箱内静置2周,进行虫体透明化,待水蒸发后进行制片。鉴定时在微分干涉显微镜(型号:Nikon-eclipse80i)下观察、鉴定、拍照,借助绘图臂绘制显微结构,利用配置软件NIS-Elements D 4.20.00测得各项形态数据。1.3 数据处理与分析
运用Excel 2019软件计算和分析小型底栖动物的类群丰度、不同站位海洋线虫的丰度。运用SPSS22软件中的单因素方差分析比较不同站位之间小型底栖动物丰度是否存在显著差异。
1.4 游咽线虫属鉴定依据
游咽线虫属(Eleutherolaimus)的主要特征:具有4+4根头刚毛。化感器圆形,下边通常具有开口。口腔圆柱形,较短,口腔壁角质化,平行。无明显食道球[14]。
游咽线虫属的种间差异主要在于:化感器相对于口腔的位置、两圈头刚毛的长度和位置、排泄孔相对于神经环的位置。在海洋线虫的分类鉴定中,会采用德曼(De Man)公式量化线虫的形态,如:a表示体长/最大体直径;b表示体长/食道长度;c表示体长/尾长;c'表示尾长/泄殖孔或肛门对应的体直径;V%表示阴门距体前端的距离/体长[15]。
2. 结果
2.1 小型底栖动物的类群组成及丰度
2022年8月份样品共鉴定出5个以上的小型底栖动物类群,分别是:海洋线虫、寡毛类、桡足类、多毛类、涡虫类和其他未鉴定类群。海洋线虫为数量最优势类群,占小型底栖动物总丰度的78.52%,其次为桡足类(19.22%),其他类群占比为2.26%。各站位小型底栖动物丰度变化趋势为:D[(492.04±416.79) 个/10 cm2]>B[(170.07±132.03) 个/10 cm2]>E[(33.05±26.76) 个/10 cm2]>C[(14.38±11.78) 个/10 cm2]>A[(4.79±2.50) 个/10 cm2]。方差分析表明,D点与A、C点小型底栖动物丰度存在显著性差异(P<0.05)。海洋线虫丰度变化趋势为D[(353.51±237.30) 个/10 cm2]>B[(166.54±128.46) 个/10 cm2]>E[(24.98±17.49) 个/10 cm2]>C[(11.61±9.28) 个/10 cm2]>A[(4.29±1.78) 个/10 cm2]。与小型底栖动物丰度站位变化趋势一致。方差分析表明,D点与A、C、E点海洋线虫丰度存在显著性差异(P<0.05)。各采样站位小型底栖动物类群丰度及占比见表2。
表 2 七股潟湖各采样站位小型底栖动物类群丰度及占比Table 2. Abundances and ratios of meiofauna groups at Qigu Lagoon类群 A B C D E 丰度/(个·10 cm−2) 占比/% 丰度/(个·10 cm−2) 占比/% 丰度/(个·10 cm−2) 占比/% 丰度/(个·10 cm−2) 占比/% 丰度/(个·10 cm−2) 占比/% 线虫 4.29±1.78 89.47 166.54±128.46 97.92 11.61±9.28 80.70 353.51±237.30 71.85 24.98±17.49 75.57 寡毛类 0±0 0 1.51±1.43 0.89 1.01±0.71 7.02 0.76±0.36 0.15 5.30±6.07 16.03 桡足类 0.50±0.71 10.53 1.01±0.71 0.59 0.76±0.36 5.26 133.99±175.21 27.23 1.01±0.71 3.05 多毛类 0±0 0 0±0 0 0.25±0.36 1.75 0±0 0 0±0 0 涡虫类 0±0 0 0.76±1.07 0.45 0.25±0.36 1.75 3.28±3.21 0.67 1.01±1.43 3.05 其他类 0±0 0 0.25±0.36 0.15 0.50±0.71 3.51 0.50±0.71 0.10 0.76±1.07 2.29 总计 4.79±2.50 100 170.07±132.03 100 14.38±11.78 100 492.04±416.79 100 33.05±26.76 100 2.2 海洋线虫群落组成及优势属
本次共鉴定七股潟湖5个站位海洋线虫491条,隶属于11科21属,海洋线虫属分布的比例及在各站位数量见表3。在不同站位之间线虫的种类组成有明显的差别,Pseudochromadora仅在A点出现;Paracanthonchus、Eleutherolaimus、Desmlaimus、Pontonema、Paracyatholaimus、Thalassomonhystera、Gnomovala仅在B点出现;Theristus、Metadesmolaimus仅在C点出现;Paraethmolaimus仅在D点出现;Molgolaimus、Ixonema仅在E点出现。
表 3 七股潟湖海洋线虫属占比及在各站位数量Table 3. Distribution proportion and quantity of marine nematode genus at various stations in Qigu Lagoon线虫属 占比% 数量/个 A B C D E Neochromadora 35.23 6 157 0 7 3 Haliplectus 34.22 0 5 0 163 0 Terschellingia 14.66 0 42 1 29 0 Monhystera 3.46 0 0 11 0 6 Daptonema 2.24 0 8 2 0 1 Paracanthonchus 1.83 0 9 0 0 0 Eleutherolaimus 1.63 0 8 0 0 0 Microlaimus 1.63 0 1 0 7 0 Paraethmolaimus 1.22 0 0 0 6 0 Parodontophora 0.61 0 1 0 2 0 Desmolaimus 0.61 0 3 0 0 0 Ptycholaimellus 0.61 1 1 1 0 0 Theristus 0.41 0 0 2 0 0 Pontonema 0.20 0 1 0 0 0 Paracyatholaimus 0.20 0 1 0 0 0 Thalassomonhystera 0.20 0 1 0 0 0 Gnomoxyala 0.20 0 1 0 0 0 Metadesmolaimus 0.20 0 0 1 0 0 Molgolaimus 0.20 0 0 0 0 1 Ixonema 0.20 0 0 0 0 1 Pseudochromadora 0.20 1 0 0 0 0 总计 100 8 239 18 214 12 以海洋线虫的丰度占比(优势度)≥5%的属作为优势属,按照优势度大小排序依次顺序为Neochromadora、Haliplectus和Terschellingia。Neochromadora在A、B、D、E点出现,优势度为35.23%;Haliplectus在B、D点出现,优势度为34.22%;Terschellingia在B、C、D点出现,优势度为14.66%。
2.3 新纪录种—拟施耐德游咽线虫(Eleutherolaimus paraschneideri)
标本采集地和生境:4个雄性和1个雌性均采集于台湾岛七股潟湖2022年8月份B站位(23.1522°N,
120.0772 °E),个体形态学测量数据见表4,显微形态结构和手绘图如图1和图2所示。表 4 拟施耐德游咽线虫的个体形态学测量数据Table 4. Individual morphological data of Eleutherolaimus paraschneideri样品 体长/μm 头直径/μm 头刚毛长/μm 亚头刚毛长/μm 口腔长/μm 口腔宽/μm 化感器距体前
端的距离/μm化感器所在
体直径/μm化感器
直径/μm♂1 2744 10.0 3.5 4.2 10.2 5.0 9.4 10.6 4.2 ♂2 2279 7.7 3.7 4.3 10.0 5.0 8.4 12.8 4.0 ♂3 2202 9.0 3.1 3.2 9.7 3.1 8.0 13.7 4.2 ♂4 2384 8.6 3.3 4.6 9.0 4.8 6.0 12.2 5.0 ♀1 2668 9.3 2.1 3.3 9.1 4.9 6.8 13.2 4.3 样品 神经环距体
前端的距离/μm神经环所在
体直径/μm食道长/μm 食道基部膨大
处的体直径/μm最大体直径/μm 排泄孔或肛门对
应的体直径/μm尾长/μm 交接器
弦长/μm交接器
弧长/μm♂1 119.0 27.3 201.8 32.3 40.0 32.9 188.6 25.2 31.2 ♂2 — — 185.6 36.0 31.7 31.3 152.3 24.4 32.3 ♂3 — — 186.6 27.7 29.4 29.9 150.4 26.0 34.0 ♂4 — — 215.0 32.0 39.5 39.0 182.8 22.0 22.4 ♀1 — — 208.6 31.3 46.0 22.6 130.2 — — 样品 引带长/μm 阴门距体
前端/μm阴门所在
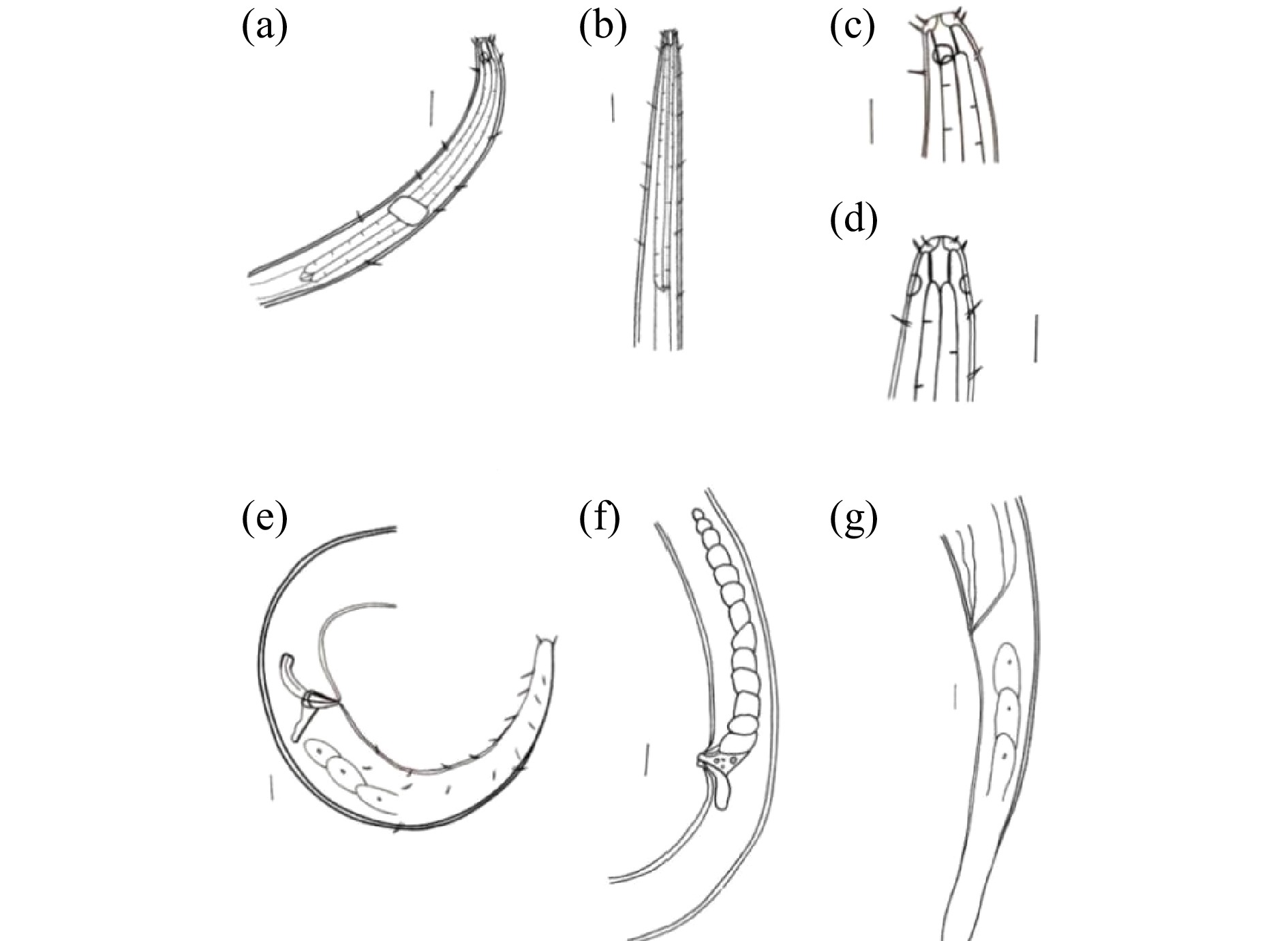
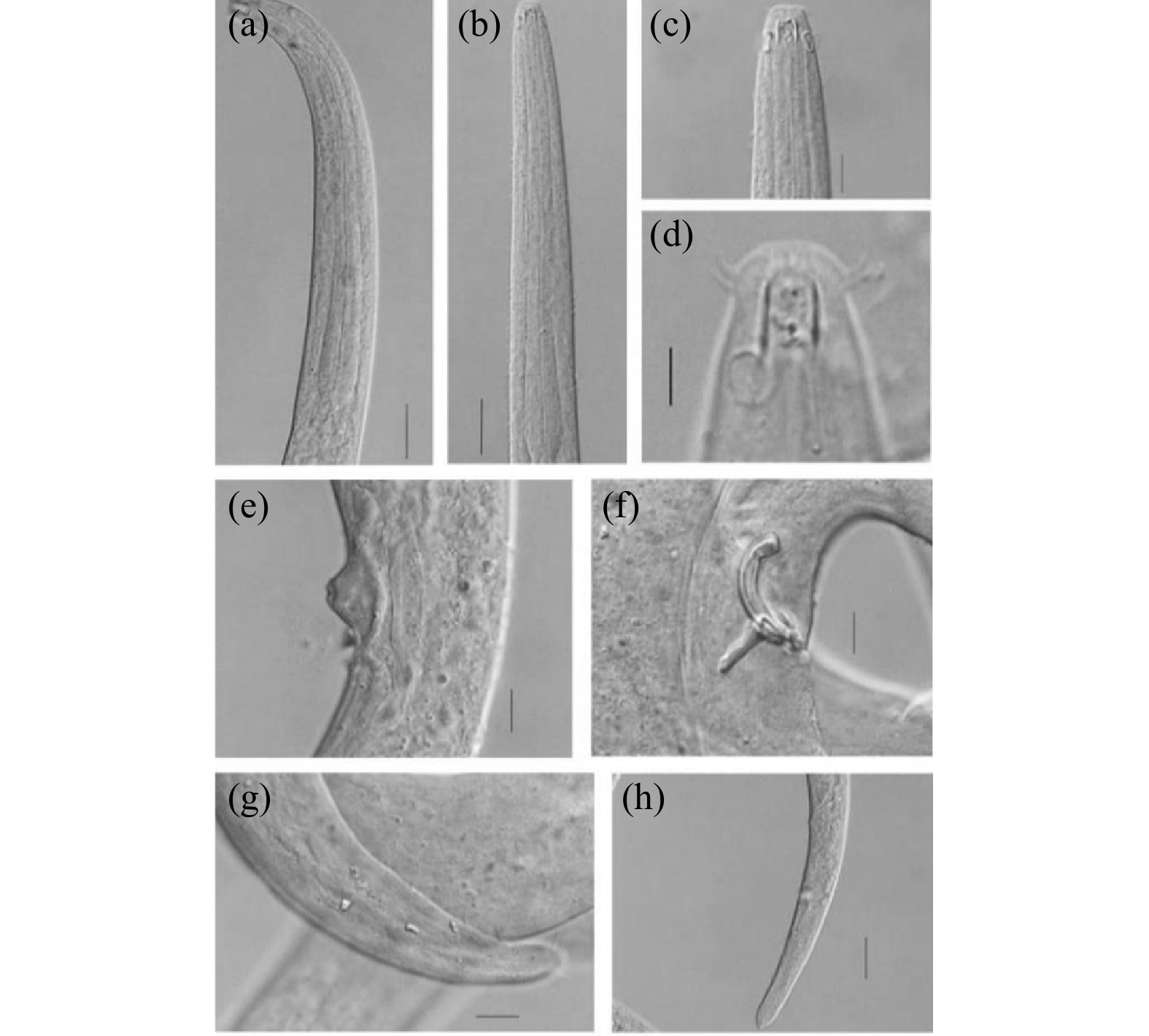
体直径/μmV% a b c c' ♂1 14.1 — — — 68.7 13.6 14.5 5.7 ♂2 17.0 — — — 72.0 12.3 15.0 4.9 ♂3 14.5 — — — 75.0 12.0 14.6 5.0 ♂4 14.0 — — — 60.4 11.1 13.0 4.7 ♀1 — 802.9 35.0 30.0 58.0 12.8 20.5 5.8 注:“—”表示该个体未测得此特征值。 ![]() 图 1 拟施耐德游咽线虫显微形态结构(a)雄性前端区域;(b)雌性前端区域;(c)雌性头部区域;(d)雄性化感器;(e)雌性阴门;(f)雄性交接器;(g)雄性尾部刚毛;(h)雌性尾部区域。比例尺:(d)为5 μm;(c)、(e)、(f)、(g)为10 μm;(a)、(b)、(h)为25 μm。Figure 1. Micromorphological structure of E. paraschneideri
图 1 拟施耐德游咽线虫显微形态结构(a)雄性前端区域;(b)雌性前端区域;(c)雌性头部区域;(d)雄性化感器;(e)雌性阴门;(f)雄性交接器;(g)雄性尾部刚毛;(h)雌性尾部区域。比例尺:(d)为5 μm;(c)、(e)、(f)、(g)为10 μm;(a)、(b)、(h)为25 μm。Figure 1. Micromorphological structure of E. paraschneideri![]() 图 2 拟施耐德游咽线虫手绘图(a)雄性前端区域;(b)雌性前端区域;(c)雄性头部区域;(d)雌性头部区域;(e)雄性交接器;(f)雌性生殖系统;(g)雌性尾部区域。比例尺:(a)、(b)为25 μm;(c)、(d)、(e)、(f)、(g)为10 μm。Figure 2. Diagram of E. paraschneideri
图 2 拟施耐德游咽线虫手绘图(a)雄性前端区域;(b)雌性前端区域;(c)雄性头部区域;(d)雌性头部区域;(e)雄性交接器;(f)雌性生殖系统;(g)雌性尾部区域。比例尺:(a)、(b)为25 μm;(c)、(d)、(e)、(f)、(g)为10 μm。Figure 2. Diagram of E. paraschneideri形态学描述:雄性身体圆柱形,向两端逐渐变细,体长为2 202~2 744 μm,最大体直径为29.4~40.0 μm(a=60.4~75.0)。虫体表皮光滑。四根头刚毛长3.1~3.7 μm,四根亚头刚毛长3.2~4.6 μm。具有稀疏分布的颈刚毛,靠近尾尖位置有成对出现的长3.0~4.2 μm的尾刚毛。口腔壁几乎平行,口腔圆柱形,长9.0~10.2 μm,宽度为3.1~5.0 μm。头部直径为8.6~10.0 μm。化感器圆形,距体前端6.0~9.4 μm,直径为4.0~5.0 μm,化感器直径占所在体直径的31%~41%,位于口腔近基部。神经环位于食道中部稍后位置,到体前端的距离占食道的59%。食道起始于口腔基部,由前向后逐渐变宽,长为185.6~215.0 μm,没有形成食道球。交接刺弯曲明显,呈弓形,弧长22.4~34.0 μm,弦长22.0~26.0 μm,近端钩状轻微膨大,远端渐尖。引带三角形的背向突起,长14.0~16.0 μm。尾长150.4~188.6 μm,为泄殖腔所在体直径的4.7~5.7倍,尾部圆锥形,末端圆钝,有3个尾腺。
雌性大部分特征与雄性相似,体长为2 668 μm,和雄性的差异在于个体较宽,最大体直径为46 μm。雌性在a值上小于雄性,为58.0,在c值上大于雄性,为20.5。阴门在身体30%处,尾部圆柱形无刚毛,尾末端略微膨大。
目前全球线虫数据库(World Database of Nematodes)[16]共记录了游咽线虫属的15个有效种,它们的形态学数据比较如表5。依据化感器和口腔位置的关系分为:化感器位于口腔近基部和口腔后面两组。七股潟湖标本化感器位于口腔近基部处属于第一组。依据口腔内无角质环、体长大小、头刚毛与亚头刚毛排列方式及长度、交接刺形态与大小,鉴定为拟施耐德游咽线虫[31]。它与模式种的区别在于,模式种a值和c值分别为90和22,七股潟湖标本a值和c值分别为60.4~75.0和13.0~15.0,均小于模式种。
表 5 游咽线虫属中有效种的形态学数据比较Table 5. Morphological data of valid species in Eleutherolaimus种名 体长/μm 头刚毛长/
μm亚头刚
毛长/μm口腔深度/
μm化感器
直径/μm化感器直径/
所在体直径交接器
弧长/μm尾长/排泄孔或肛门
对应的体直径a E. amasi[17] ♂ 2200 ~2737 3 9 7 8 62% 28~35 5.2~7.3 88~95 ♀ 2240 ~3034 3.5 7~9 6.5~7.0 50%~51% 5.8~8.4 78~97 E. chesapeakiensis[18] ♂ 3000 ~3700 — — — — 45% 35 4.1~4.3 91~100 E. doliolum[19] ♀ 1850 4 14 10 7~8 55%~60% — 7 74 E. hopperi[20] ♂ 1270 ~1740 4~5 9~15 8~9 5.5~7.0 60%~80% 20~22 4.2~4.3 58~82 ♀ 1690 ~2230 9~15 5~6 42%~67% — 4.7~6.0 63~103 E. inglisi[21] ♂ 1570 ~1950 3~4 5 12~14 5 55%~60% 22~25 3.7~4.7 53~66 ♀ 1500 ~2190 4~5 6 — — 3.7~5.0 47~70 E. iniquisetosus[22] ♂ 2142 4 13~15 15.0~17.5 — 47% 34 5.1 60 ♀ 2060 ~2786 2~3 11~12 7~8 50%~55% — 5.7~7.0 57~72 E. longus[23] ♂ 1670 4 — 6 7 50% 32 4.04 55 E. minutus[24] ♂940~ 1110 2.0~2.5 1.5~2.0 3.5~4.0 4~5 50%~63% 14~16 7.45~8.35 72~93 ♀940~ 1139 2 1.5 — 4~5 47%~62% — 6.2~8.2 67~89 E. nutus[25] ♂ 2370 4~5 11~13 13 6.5 50% 33 5.94 61 ♀ 3157 5.5 — — — 65 E. obtusicaudatus[26] ♂ 4070 ~6800 5 6.0~6.5 8.5 6.0~6.5 33% 42~52 3.4~5.5 88~128 ♀ 2753 ~6800 — 6.5 25%~33% — 3.5~5.2 106~128 E. parasabulicolus[27] ♂ 2767 ~3040 5 5 22 10 35% 67~71 2.3~3.3 36~43 ♀ 3400 — — — 9~10 50%~60% — — — E. sabulicolius[28] ♂ 4100 ~4300 4 10~15 15~20 7 40% 33~37 5.5~6.0 52~86 E. schneideri[29] ♂ 1387 ~2349 2.0~3.5 5~7 8~12 6~7 50%~55% 21~27 4.2~8.0 82~99 ♀ 1251 ~1828 — — — 5.0~6.5 45%~50% — 3.7~6.9 57~94 E. stenosoma[30] ♂ 1558 ~2500 2.0~2.5 6.5~7.5 5~7 3.3~5.0 43%~44% 21.5~36.0 6.5 70~81 ♀ 1670 ~2500 1~3 3.5~5.0 6.5~8.0 3.7~5.0 42%~50% — 7.9~8.0 56~70 E. paraschneideri[31] ♂ 2620 3~4 3~4 — 5 42% 21 4.6 90 ♀ 2816 3~5 3~5 — 5 38% — 5 85 E. paraschneideri
(本研究)♂ 2202 ~2744 3.1~3.7 3.2~4.6 9.0~10.2 4~5 31%~41% 22.4~34.0 4.9~5.7 60.4~75.0 ♀ 2668 2.1 3.3 9.1 4.3 33% — 5.8 58 注:“—”表示数据缺失。 七股潟湖的标本与E. chesapeakiensis、E. schneideri和E. inglisi 3个种也相似。与E. chesapeakiensis的区别在于食道部分具有刚毛,交接刺形态不同[18]。与E. schneideri的区别在于除食道、尾部外不具有体刚毛,体长相差较大(
2202 ~2744 μm vs.1387 ~2349 μm),交接刺形态不同[29]。在孟加拉孙德尔本斯红树林被首次发现的E. inglisi[21]口腔内有角质环,本研究的标本无角质环,化感器直径占所在体直径的比值相差也较大(31%~41% vs. 55%~60%),引带三角形的背向突起长不同(14~16 μm vs. 17~19 μm)。3. 讨论
砂质沙滩生物周期性地暴露于空气中,频繁的温差波动、盐度变化以及波浪和海流的侵袭,是小型底栖动物丰度数量少的因素[32-33]。七股潟湖为半封闭的沙洲型潟湖,处在淡水和海水交汇区域,同时,受涨落潮影响,环境条件变动剧烈,属于高度动态的生态系统[13]。沿岸流输沙往南,加上台风波浪侵蚀及暴潮与水流上溯的影响,导致七股潟湖沙洲存在侵蚀后缩、潟湖的严重陆化[12]。Alongi研究澳大利亚大堡礁戴维斯礁海洋线虫的种群结构,发现未受保护的浅潟湖区域沉积物受到强烈的波浪作用比其他礁区海洋线虫丰度低,一般小于60 个/10 cm2[34]。本研究小型底栖动物的丰度介于(4.79±2.50)~(492.04±416.79) 个/10 cm2之间,海洋线虫的丰度介于(4.29±1.78)~(353.51±237.30) 个/10 cm2之间,A、C、E点的丰度小于60 个/10 cm2。
根据邱英哲等对七股潟湖底质环境调查结果显示,A点和E点底质粒径大于B和C点,大颗粒底质保水性不佳,有机质积累量少,导致底栖动物数量减少[13]。蔡立哲等对台湾淡水河口潮间带小型底栖动物组成研究发现,沙滩的丰度远低于红树林和有芦苇的红树林区域,小型底栖动物的丰度介于(4.67±2.08)~(294.00±180.40) 个/10 cm2之间[8]。王玥等在夏季广西北海金海湾红树林湿地采样发现,光滩的小型底栖动物数量低于红树林区域,小型底栖动物的丰度介于(6.07±1.23)~(200.25±31.75) 个/10 cm2之间,海洋线虫的丰度介于(4.16±1.70)~(195.23±30.80) 个/10 cm2之间[35]。本研究中,A、E点沙洲小型底栖动物和海洋线虫丰度低分别为(4.79±2.50) 个/10 cm2和(33.05±26.76) 个/10 cm2,特别是A点,受到上游输沙、海水汇入与牡蛎养殖水排出等因素的干扰[13]。另外,旅游者的踩踏会直接造成沉积物溶氧量含量降低[36],导致小型底栖动物丰度及分布的变化。七股潟湖也是台湾重要的旅游景点[12],这些因素综合起来可能是A点小型底栖动物和海洋线虫丰度为最低的原因。
有研究发现,在有丰富有机质输入的沙滩,小型底栖动物和海洋线虫数量丰富[37]。七股潟湖各站位有机质含量从高到低的顺序依次为:B(5.53±6.26)%>D(4.05±3.86)%>C(3.75±1.43)%>A(2.93±1.01)%>E(2.80±3.07)%[13]。B和D点有机质含量相对较高,可能也是小型底栖和海洋线虫丰度大于A、C、E点的原因。据邱英哲等[13]对七股潟湖多毛类群与环境因子之间的关系调查发现,C点底质以细密粉砂泥为主,低溶氧环境,多毛类的丰度为七股潟湖潮间带最低。本研究C点海洋线虫丰度为(14.38±11.78) 个/10 cm2,较低的原因可能与有机质含量低、沉积物类型为细密粉砂泥有关。
Ansari等在亚洲最大沿海潟湖印度奇利卡观察到海洋线虫13科32属64种,其优势属为Oncholaimellus、Viscosia、Dorylaimopsis、Sabatieria、Daptonema和Terschellingia[38]。本研究中Terschellingia是优势属,分布在B、C、D点。本研究与宋慧丽等[39]在冬季大连金沙滩发现的海洋线虫优势属仅有Neochromadora相同。肖月平等在金门岛红树林湿地发现的Daptonema为优势属[40],本研究发现的Daptonema属在泥滩和沙洲均有出现。周仁桂等在深圳红树林发现海洋线虫Haliplectus属的两个新种[41],Haliplectus作为本研究的优势属在泥滩中出现。不同线虫属对环境适应力存在一定差异,从而使其在各种生境中海洋线虫的优势程度不同,综上所述,Terschellingia、Neochromadora、Daptonema、Haliplectus不仅在泥滩中为优势属,在红树林中也具有较高丰度。
游咽线虫属隶属于条线虫科(Linhomoeidae),单宫目(Monhysterida),由Filipjev在1922年建立,并将Eleutherolaimus longus作为模式种[23]。目前研究发现该属物种分布较广,在北冰洋弗拉姆海峡的莫洛伊深海
5000 m处[24]、埃姆斯河口泥滩[28]、新西兰坎伯恩附近的帕瓦塔哈努伊湾的砂质潮间带[31]以及孟加拉孙德尔本斯红树林[20-21]中均有报道。国内,黄勇等在东海记录了E. chesapeakensis的出现[14];该属以优势属在莱州湾出现[42];福建同安湾红树林也有出现[43]。4. 结论
本研究共在台湾岛南部七股潟湖沉积物中鉴定出5个以上的小型底栖动物类群;小型底栖动物的丰度介于(4.79±2.50)~(492.04±416.79) 个/10 cm2之间,其中靠近七股潟湖北潮口A站位的小型底栖动物和海洋线虫丰度为最低;海洋线虫群落隶属于11科21属,优势属为Neochromadora、Haliplectus和Terschellingia;发现国内新纪录种—拟施耐德游咽线虫(Eleutherolaimus paraschneideri),主要特征为:口腔内无角质环,化感器位于口腔近基部。交接器呈弓形,近端轻微肿胀,引带具三角形的背向突起。
-
![]()
图 1 拟施耐德游咽线虫显微形态结构
(a)雄性前端区域;(b)雌性前端区域;(c)雌性头部区域;(d)雄性化感器;(e)雌性阴门;(f)雄性交接器;(g)雄性尾部刚毛;(h)雌性尾部区域。比例尺:(d)为5 μm;(c)、(e)、(f)、(g)为10 μm;(a)、(b)、(h)为25 μm。
Figure 1. Micromorphological structure of E. paraschneideri
![]()
图 2 拟施耐德游咽线虫手绘图
(a)雄性前端区域;(b)雌性前端区域;(c)雄性头部区域;(d)雌性头部区域;(e)雄性交接器;(f)雌性生殖系统;(g)雌性尾部区域。比例尺:(a)、(b)为25 μm;(c)、(d)、(e)、(f)、(g)为10 μm。
Figure 2. Diagram of E. paraschneideri
表 1 七股潟湖采样站位信息及其生境
Table 1 Sampling station information and habitat of Qigu Lagoon
采样站位 经度 纬度 生境 A 120.0746 °E23.1725 °N沙洲 B 120.0772 °E23.1522 °N泥滩 C 120.0871 °E23.1307 °N红树林 D 120.0881 °E23.1174 °N泥滩 E 120.0397 °E23.1057 °N沙洲  下载: 导出CSV
下载: 导出CSV
表 2 七股潟湖各采样站位小型底栖动物类群丰度及占比
Table 2 Abundances and ratios of meiofauna groups at Qigu Lagoon
类群 A B C D E 丰度/(个·10 cm−2) 占比/% 丰度/(个·10 cm−2) 占比/% 丰度/(个·10 cm−2) 占比/% 丰度/(个·10 cm−2) 占比/% 丰度/(个·10 cm−2) 占比/% 线虫 4.29±1.78 89.47 166.54±128.46 97.92 11.61±9.28 80.70 353.51±237.30 71.85 24.98±17.49 75.57 寡毛类 0±0 0 1.51±1.43 0.89 1.01±0.71 7.02 0.76±0.36 0.15 5.30±6.07 16.03 桡足类 0.50±0.71 10.53 1.01±0.71 0.59 0.76±0.36 5.26 133.99±175.21 27.23 1.01±0.71 3.05 多毛类 0±0 0 0±0 0 0.25±0.36 1.75 0±0 0 0±0 0 涡虫类 0±0 0 0.76±1.07 0.45 0.25±0.36 1.75 3.28±3.21 0.67 1.01±1.43 3.05 其他类 0±0 0 0.25±0.36 0.15 0.50±0.71 3.51 0.50±0.71 0.10 0.76±1.07 2.29 总计 4.79±2.50 100 170.07±132.03 100 14.38±11.78 100 492.04±416.79 100 33.05±26.76 100
下载: 导出CSV
表 3 七股潟湖海洋线虫属占比及在各站位数量
Table 3 Distribution proportion and quantity of marine nematode genus at various stations in Qigu Lagoon
线虫属 占比% 数量/个 A B C D E Neochromadora 35.23 6 157 0 7 3 Haliplectus 34.22 0 5 0 163 0 Terschellingia 14.66 0 42 1 29 0 Monhystera 3.46 0 0 11 0 6 Daptonema 2.24 0 8 2 0 1 Paracanthonchus 1.83 0 9 0 0 0 Eleutherolaimus 1.63 0 8 0 0 0 Microlaimus 1.63 0 1 0 7 0 Paraethmolaimus 1.22 0 0 0 6 0 Parodontophora 0.61 0 1 0 2 0 Desmolaimus 0.61 0 3 0 0 0 Ptycholaimellus 0.61 1 1 1 0 0 Theristus 0.41 0 0 2 0 0 Pontonema 0.20 0 1 0 0 0 Paracyatholaimus 0.20 0 1 0 0 0 Thalassomonhystera 0.20 0 1 0 0 0 Gnomoxyala 0.20 0 1 0 0 0 Metadesmolaimus 0.20 0 0 1 0 0 Molgolaimus 0.20 0 0 0 0 1 Ixonema 0.20 0 0 0 0 1 Pseudochromadora 0.20 1 0 0 0 0 总计 100 8 239 18 214 12
下载: 导出CSV
表 4 拟施耐德游咽线虫的个体形态学测量数据
Table 4 Individual morphological data of Eleutherolaimus paraschneideri
样品 体长/μm 头直径/μm 头刚毛长/μm 亚头刚毛长/μm 口腔长/μm 口腔宽/μm 化感器距体前
端的距离/μm化感器所在
体直径/μm化感器
直径/μm♂1 2744 10.0 3.5 4.2 10.2 5.0 9.4 10.6 4.2 ♂2 2279 7.7 3.7 4.3 10.0 5.0 8.4 12.8 4.0 ♂3 2202 9.0 3.1 3.2 9.7 3.1 8.0 13.7 4.2 ♂4 2384 8.6 3.3 4.6 9.0 4.8 6.0 12.2 5.0 ♀1 2668 9.3 2.1 3.3 9.1 4.9 6.8 13.2 4.3 样品 神经环距体
前端的距离/μm神经环所在
体直径/μm食道长/μm 食道基部膨大
处的体直径/μm最大体直径/μm 排泄孔或肛门对
应的体直径/μm尾长/μm 交接器
弦长/μm交接器
弧长/μm♂1 119.0 27.3 201.8 32.3 40.0 32.9 188.6 25.2 31.2 ♂2 — — 185.6 36.0 31.7 31.3 152.3 24.4 32.3 ♂3 — — 186.6 27.7 29.4 29.9 150.4 26.0 34.0 ♂4 — — 215.0 32.0 39.5 39.0 182.8 22.0 22.4 ♀1 — — 208.6 31.3 46.0 22.6 130.2 — — 样品 引带长/μm 阴门距体
前端/μm阴门所在
体直径/μmV% a b c c' ♂1 14.1 — — — 68.7 13.6 14.5 5.7 ♂2 17.0 — — — 72.0 12.3 15.0 4.9 ♂3 14.5 — — — 75.0 12.0 14.6 5.0 ♂4 14.0 — — — 60.4 11.1 13.0 4.7 ♀1 — 802.9 35.0 30.0 58.0 12.8 20.5 5.8 注:“—”表示该个体未测得此特征值。
下载: 导出CSV
表 5 游咽线虫属中有效种的形态学数据比较
Table 5 Morphological data of valid species in Eleutherolaimus
种名 体长/μm 头刚毛长/
μm亚头刚
毛长/μm口腔深度/
μm化感器
直径/μm化感器直径/
所在体直径交接器
弧长/μm尾长/排泄孔或肛门
对应的体直径a E. amasi[17] ♂ 2200 ~2737 3 9 7 8 62% 28~35 5.2~7.3 88~95 ♀ 2240 ~3034 3.5 7~9 6.5~7.0 50%~51% 5.8~8.4 78~97 E. chesapeakiensis[18] ♂ 3000 ~3700 — — — — 45% 35 4.1~4.3 91~100 E. doliolum[19] ♀ 1850 4 14 10 7~8 55%~60% — 7 74 E. hopperi[20] ♂ 1270 ~1740 4~5 9~15 8~9 5.5~7.0 60%~80% 20~22 4.2~4.3 58~82 ♀ 1690 ~2230 9~15 5~6 42%~67% — 4.7~6.0 63~103 E. inglisi[21] ♂ 1570 ~1950 3~4 5 12~14 5 55%~60% 22~25 3.7~4.7 53~66 ♀ 1500 ~2190 4~5 6 — — 3.7~5.0 47~70 E. iniquisetosus[22] ♂ 2142 4 13~15 15.0~17.5 — 47% 34 5.1 60 ♀ 2060 ~2786 2~3 11~12 7~8 50%~55% — 5.7~7.0 57~72 E. longus[23] ♂ 1670 4 — 6 7 50% 32 4.04 55 E. minutus[24] ♂940~ 1110 2.0~2.5 1.5~2.0 3.5~4.0 4~5 50%~63% 14~16 7.45~8.35 72~93 ♀940~ 1139 2 1.5 — 4~5 47%~62% — 6.2~8.2 67~89 E. nutus[25] ♂ 2370 4~5 11~13 13 6.5 50% 33 5.94 61 ♀ 3157 5.5 — — — 65 E. obtusicaudatus[26] ♂ 4070 ~6800 5 6.0~6.5 8.5 6.0~6.5 33% 42~52 3.4~5.5 88~128 ♀ 2753 ~6800 — 6.5 25%~33% — 3.5~5.2 106~128 E. parasabulicolus[27] ♂ 2767 ~3040 5 5 22 10 35% 67~71 2.3~3.3 36~43 ♀ 3400 — — — 9~10 50%~60% — — — E. sabulicolius[28] ♂ 4100 ~4300 4 10~15 15~20 7 40% 33~37 5.5~6.0 52~86 E. schneideri[29] ♂ 1387 ~2349 2.0~3.5 5~7 8~12 6~7 50%~55% 21~27 4.2~8.0 82~99 ♀ 1251 ~1828 — — — 5.0~6.5 45%~50% — 3.7~6.9 57~94 E. stenosoma[30] ♂ 1558 ~2500 2.0~2.5 6.5~7.5 5~7 3.3~5.0 43%~44% 21.5~36.0 6.5 70~81 ♀ 1670 ~2500 1~3 3.5~5.0 6.5~8.0 3.7~5.0 42%~50% — 7.9~8.0 56~70 E. paraschneideri[31] ♂ 2620 3~4 3~4 — 5 42% 21 4.6 90 ♀ 2816 3~5 3~5 — 5 38% — 5 85 E. paraschneideri
(本研究)♂ 2202 ~2744 3.1~3.7 3.2~4.6 9.0~10.2 4~5 31%~41% 22.4~34.0 4.9~5.7 60.4~75.0 ♀ 2668 2.1 3.3 9.1 4.3 33% — 5.8 58 注:“—”表示数据缺失。
下载: 导出CSV
-
[1] 张志南,周红. 国际小型底栖生物研究的某些进展[J]. 中国海洋大学学报(自然科学版),2004,34(5):799-806. ZHANG Z N,ZHOU H. Some progress on the study of meiofauna[J]. Periodical of Ocean University of China,2004,34(5):799-806.
[2] 杜永芬,徐奎栋,类彦立. 海洋微型和小型底栖生物相互作用研究综述[J]. 海洋科学集刊,2009(1):163-173. DU Y F,XU K D,LEI Y L. Research progress in the interactions of marine micro-and meiobenthos[J]. Studia Marina Sinica,2009(1):163-173.
[3] 林俊辉,郑凤武. 泉州湾及其附近海域底栖多毛类生态的初步研究[J]. 台湾海峡,2007,26(2):281-288. LIN J H,ZHENG F W. Preliminary study on ecology of benthic Polychaeta in Quanzhou Bay and its adjacent waters[J]. Journal of Oceanography in Taiwan Strait,2007,26(2):281-288.
[4] 徐奎栋,洪华生,宋微波,等. 台湾海峡的砂壳纤毛虫研究(纤毛动物门:砂壳亚目)[J]. 动物分类学报,2001,26(4):454-466. DOI: 10.3969/j.issn.1000-0739.2001.04.006 XU K D,HONG H S,SONG W B,et al. Studies on tintinnine ciliates in the Taiwan Strait (Ciliophora:Tintinnina)[J]. Acta Zootaxonomica Sinica,2001,26(4):454-466. DOI: 10.3969/j.issn.1000-0739.2001.04.006
[5] 方少华,吕小梅,张跃平,等. 台湾海峡小型底栖生物数量的量分布[J]. 海洋学报,2000,22(6):136-140. FANG S H,LV X M,ZHANG Y P,et al. Distribution abundance of meiobenthos in the Taiwan Strait[J]. Acta Oceanologica Sinica,2000,22(6):136-140.
[6] 林元烧,罗文新,曹文清,等. 台湾海峡异养性鞭毛虫生态研究I:1997年夏季南部海域鞭毛虫丰度及生物量分布[J]. 厦门大学学报(自然科学版),2001,40(3):798-803. LIN Y S,LUO W X,CAO W Q,et al. Studies on heterotrophic nanoflagellates in Taiwan strait Ⅰ:distribution of abundance and biomass of HNF in southern waters,summer of 1997[J]. Journal of Xiamen University (Natural Science),2001,40(3):798-803.
[7] 王彦国,林茂,项鹏,等. 台湾海峡中北部海域春季小型底栖生物丰度和生物量[J]. 台湾海峡,2010,29(3):373-379. WANG Y G,LIN M,XIANG P,et al. Abundance and biomass of meiobenthos in central and northern Taiwan Strait during spring[J]. Journal of Oceanography in Taiwan Strait,2010,29(3):373-379.
[8] CAI L Z,FU S J,ZHOU X P,et al. Benthic meiofauna with emphasis on nematode assemblage response to environmental variation in the intertidal zone of the Danshuei River Estuary,northwest Taiwan[J]. Ecological Research,2020,35(5):857-870. DOI: 10.1111/1440-1703.12159
[9] XIAO Y P,GUO Y Q. Two new and one known free-living marine nematode species of the family Axonolaimidae from coastal wetlands in western Taiwan[J]. Regional Studies in Marine Science,2023,60:1-13.
[10] XIAO Y P,GUO Y Q. Characterization of two new free-living marine nematode species of the genus Trissonchulus from the mangrove wetlands of western Taiwan Island[J]. Cahiers de Biologie Marine,2023,64:235-246.
[11] KJERFVE B. Chapter 1 coastal lagoons[M]//KJERFVE B. Coastal lagoon processes. Amsterdam:Elsevier,1994:1-8.
[12] 吴盈志,刘景毅,黄煌辉. 七股潟湖沙洲地形变迁之研究[J]. 海洋工程学刊,2013(13):367-391. WU Y Z,LIU J Y,HUANG H H. The morphological study on Qigu Lagoon[J]. Journal of Coastal and Ocean Engineering,2013(13):367-391.
[13] 邱英哲,沈子耘,黄家勤,等. 七股潟湖潮间带底栖多毛类群聚及其与环境因子之关系[J]. 台湾生物多样性研究,2011,13(2): 135-151. QIU Y Z,SHEN Z Y,HUANG J Q,et al. Abundance and species diversity of benthic polychaetes in relation to sediment environmental variables in the intertidal zone of Qigu Lagoon, Taiwan[J]. TW J. of Biodivers,2011,13(2): 135-151.
[14] 黄勇,郭玉清,翟红秀. 东海自由生活海洋线虫分类研究[M]. 北京:科学出版社,2022:168-242. HUANG Y,GUO Y Q,ZHAI H X. Classification of free-living marine nematodes in the East China Sea[M]. Beijing:Science Press,2022:168-242.
[15] PLATT H M,WARWICK R M. Free-living marine nematodes. Part Ⅱ:British Chromadorids[M]. London:Synopses of the British Fauna(New Series),1988.
[16] Nemys. World database of nematodes[DB/OL].[2023-09-27]. https://nemys.ugent.be.
[17] BOUWMAN L A. A survey of nematodes from the Ems Estuary Part I:Systematics.[J]. Zool. Jb. (Syst. ),1981,108:335-385.
[18] TIMM R W. A survey of the marine nematodes of Chesapeake Bay,Maryland[D]. Washington, D.C.: The Catholic University of America,1954.
[19] WIESER W. Free-living nematodes and other small invertebrates of Puget Sound beaches[J]. University of Washington Publications in Biology,1959,19:1-179.
[20] TIMM R W. New marine nematodes of the family Linhomoeidae from East Pakistan[J]. Proceedings of the Pakistan Academy of Science,1967,4(1):15-22.
[21] TIMM R W. Some estuarine nematodes from the Sunderbans[J]. Proceedings of the Pakistan Academy of Science,1967,4:1-3.
[22] SCHUURMANS STEKHOVEN J H. Freeliving marine Nemas of the Belgian coast. I. and II[J]. Mém. Mus. R. Hist. Nat. Belg,1935,72:1-36.
[23] FILIPJEV I N. Encore sur les Nématodes libres de la mer Noire[J]. Tr. Stravrop. Skh. Inst. Zool,1922,1:83-184.
[24] TCHESUNOV A V,MILJUTIN D M. Three new free-living nematode species (Monhysterida) from the Arctic abyss,with revision of the genus Eleutherolaimus Filipjev,1922 (Linhomoeidae)[J]. Russian Journal of Nematology,2006,14(1):57-75.
[25] GERLACH S A. Freilebende Nematoden aus dem Roten Meer[J]. Kieler Meeresforsch,1964,20:18-34.
[26] ALLGEN C A. West American marine nematodes[J]. Vidensk. Medd. fra Dansk naturh. Foren,1947,110:65-219.
[27] ALLGEN C A. Die freilebenden Nematoden des Öresunds[J]. Capita Zoologica,1935,6(3):1-192.
[28] SCHULZ E. Beiträge zur Kenntnis mariner Nematoden aus der Kieler Bucht[J]. Zool. Jahrb. Jena (Syst.),1932,62:331-430.
[29] TURPEENNIEMI T A. Four new nematode species from the Bothnian bay,northern Baltic Sea,with a redescription of Microlaimus globiceps de man,1880 (Nematoda)[J]. Nematologica,1997,43(1):31-58. DOI: 10.1163/004725997X00034
[30] DE MAN J G. Sur quelques espèces nouvelles ou peu connues de nématodes libres vivant sur les côtes de la Zélande[J]. Tijdschrift Der Nederlandsche Dierkundige Vereeniging,1970,10:227-244.
[31] LEDUC D,ZHAO Z Q. The marine biota of Aotearoa New Zealand[M]. Wellington: National Institute of Water and Atmospheric Research,2023:135-212.
[32] 陈玉珍,郭玉清,刘爱原. 福建省4个主要岛屿沙滩小型底栖动物的丰度与分布差异[J]. 厦门大学学报(自然科学版),2020,59(2):254-262. CHEN Y Z,GUO Y Q,LIU A Y. Differences of meiofaunal abundances and distributions in the sandy beaches of four main islands in Fujian Province[J]. Journal of Xiamen University (Natural Science),2020,59(2):254-262.
[33] 陈玉珍,郭玉清,刘爱原. 福建海坛岛长江澳沙滩自由生活海洋线虫群落研究[J]. 生态学报,2019,39(7):2573-2582. CHEN Y Z,GUO Y Q,LIU A Y. Community structures of free-living marine nematodes on sandy beach of Changjiangao (Haitan Island,Fujian Province)[J]. Acta Ecologica Sinica,2019,39(7):2573-2582.
[34] ALONGI D M. Population structure and trophic composition of the free-living nematodes inhabiting carbonate sands of Davies Reef,Great Barrier Reef,Australia[J]. Marine and Freshwater Research,1986,37(5):609. DOI: 10.1071/MF9860609
[35] 王玥,庞小鹏,郭玉清. 广西北海金海湾红树林湿地海洋线虫群落研究初探[J]. 海洋与湖沼,2020,51(3):583-590. DOI: 10.11693/hyhz20191100239 WANG Y,PANG X P,GUO Y Q. Study on marine nematodes community in Jinhai Bay mangrove wetland,Beihai,Guangxi[J]. Oceanologia et Limnologia Sinica,2020,51(3):583-590. DOI: 10.11693/hyhz20191100239
[36] 李佳,华尔,张志南. 青岛砂质潮间带小型底栖动物分布及季节动态[J]. 应用生态学报,2012,23(12):3458-3466. LI J,HUA E,ZHANG Z N. Distribution and seasonal dynamics of meiofauna in intertidal zone of Qingdao sandy beaches,Shandong Province of East China[J]. Chinese Journal of Applied Ecology,2012,23(12):3458-3466.
[37] MENN I. Beach morphology and food web structure:comparison of an eroding and an accreting sandy shore in the North Sea[J]. Helgoland Marine Research,2002,56(3):177-189. DOI: 10.1007/s10152-002-0108-0
[38] ANSARI K G M T,PATTNAIK A K,RASTOGI G,et al. An inventory of free-living marine nematodes from Asia’s largest coastal lagoon,Chilika,India[J]. Wetlands Ecology and Management,2015,23(5):881-890. DOI: 10.1007/s11273-015-9426-2
[39] 宋慧丽,慕芳红,孙燕,等. 冬季大连砂质潮间带自由生活海洋线虫群落结构及多样性的比较[J]. 海洋学报,2021,43(8):139-151. SONG H L,MU F H,SUN Y,et al. Comparison of community structure and diversity of free-living marine nematodes in the sandy intertidal zone of Dalian in winter[J]. Haiyang Xuebao,2021,43(8):139-151.
[40] 肖月平,郭玉清,施宜佳. 金门岛红树林湿地海洋线虫群落结构的初步探究[J]. 应用海洋学学报,2023,42(3):402-408. XIAO Y P,GUO Y Q,SHIH Y J. Community structures of marine nematodes in Jinmen mangrove wetland[J]. Journal of Applied Oceanography,2023,42(3):402-408.
[41] ZHOU R G,CHEN Y Z,SHIH Y J,et al. Two new free-living marine nematode species of the genus Haliplectus Cobb,1913 (Haliplectidae) from mangroves of Shenzhen,Guangdong Province,China[J]. Journal of Marine Science and Engineering,2022,10(11):1694. DOI: 10.3390/jmse10111694
[42] 孙燕. 莱州湾小型底栖生物生态和自由生活线虫分类研究[D]. 聊城:聊城大学,2018. SUN Y. Study on ecology of meiobenthos and classification of free-living nematodes in Laizhou Bay[D]. Liaocheng:Liaocheng University,2018.
[43] 曾佳丽. 福建同安湾红树林湿地自由生活线虫的分类研究[D]. 厦门:厦门大学,2017. ZENG J L. Classification of the free living nematodes in Tong’an Bay mangrove wetland,Fujian[D]. Xiamen:Xiamen University,2017.
计量
- 文章访问数: 13
- HTML全文浏览量: 3
- PDF下载量: 0
