
Spatial pattern and connectivity of marine ecological protection in the Yangtze River Estuary based on maritime management
-
摘要:
生态保护空间格局及其连通性对保护生物物种和维持局域生物多样性具有重要作用。本研究以长江口海域空间利用管理模式为基础,利用形态学空间格局分析(Morphological Spatial Pattern Analysis, MSPA)和景观连通性模型,研究长江口海域生境特征,识别重要生态源地及其连通性,并分析其与现有保护格局的匹配性,以期为长江口海域生态系统整体保护、系统修复、综合治理提供支持。研究结果表明:长江口海域15块保护空间中,识别重要生态源地12个,总面积约为
3080 km2,占上海市海洋功能区划面积的28.6%。重要源地主要分布于长江口南支及附近海域。连接12个生态源地的生态廊道23条。本研究识别的长江口门内生态源地空间与上海市最新公布的海洋生态保护红线空间匹配性较好,但连接南支和北支口门附近的重要保护空间不足。Abstract:The ecological space protection pattern and its connectivity play an important role in maintaining local biodiversity. This study is based on the spatial utilization management of theYangtze River Estuary (YRE) in Shanghai, and uses morphological spatial pattern analysis (MPSA) and landscape connectivity models to study the habitat characteristics of the YRE based on the spatial management model of the sea area. It identifies important ecological source areas and their connectivity and analyzes their compatibility with the existing protection pattern in order to contribute to the integrated protection of the YRE. Results indicate that there are 12 main ecological source areas in the YRE, with a total area of approximately
3080 km2 accounting for 28.6% of the areas of Shanghai’s Marine Functional Zoning. The important source areas are mainly located in the southern branch of the YRE and the nearby sea area with 23 ecological corridors connecting 12 ecological source areas. The ecological source space identified in this study matches well with the latest marine ecological protection red line space announced. However, the important protection space is insufficient outside the river mouth, especially in important protection space near the southern and northern branches.-
Keywords:
- maritime management /
- ecological protect area /
- pattern /
- connectivity /
- the Yangtze River Estuary
-
维护区域生态系统及其生态过程的稳定是生态保护网络构建的重要目标[1]。至今,国内外学者针对如何构建栖息地网络开展了大量研究[2-4],生态网络构建作为一种重要的规划技术手段成为近年来相关研究的热点[5-8]。但这一技术方法很少在水生生态系统中应用。河口海域主要以河口径流和海水相互稀释过程中形成的淡水-咸淡水-咸水的连续水体组成,其生境的多样性为许多河口鱼类提供了重要生境,如索饵、育幼等[9]。特别是河口区与海洋连通性显著影响其关键物理化学变量,生境的异质性可以调节该区域生态系统之间的相互连接程度[10]。在潮汐等动力作用下,生境异质性往往受河口径流与海水层化、混合等过程影响[11],如Friesen等研究表明近岸海水温度变化影响底栖生物连通性[12]。连通的界限取决于研究区域以及所考虑的物种[13]。
然而,河口海域是人类开发活动最为频繁的区域之一,其生境连通性不仅受自然环境的影响,人类活动及其利用模式也对其产生显著的重塑作用。自2011年国家实施海洋功能区划以来,我国近岸海域各类开发活动均需符合海洋功能区划空间管控要求,10年来基于海洋功能区划的空间管理,深度影响了海洋生境分布,被划分为开发利用空间的海域,其作为物种生境的功能基本丧失,而区划为保护空间的海域被赋予区域生态源地的功能。然而这些生态源地的作用如何,各源地间是否具有有效的连通性,对于这一问题至今仍鲜有研究进行探讨。本研究通过构建基于长江口生态源地识别和连通性评估的技术方法,并评估空间利用模式对生境的重塑作用,以期为我国“十四五”以来基于国土空间规划的海域利用管理新模式提供技术支持。
1. 研究区概况和数据来源
1.1 研究区概况
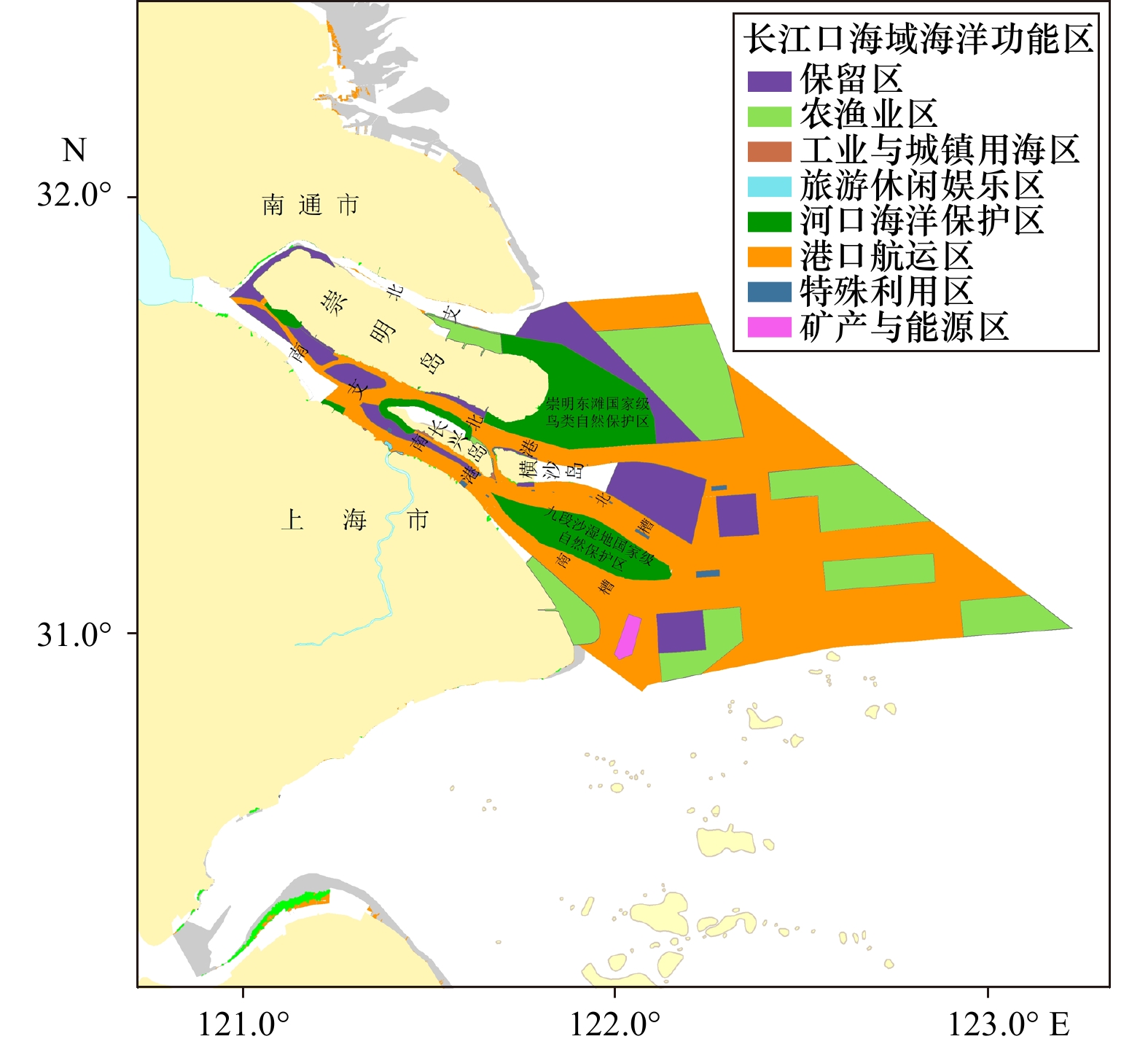
研究区位于上海市长江口海域,分布有国家级自然保护地、水源地保护区、自然保护红线等各类保护空间,是上海市构建生态屏障的重要区域。该海域主要由长江口和其毗邻海域构成,面积约为
8000 km2。长江口为三级分汊、四口入海的三角洲河口,其河槽被崇明岛分为南支和北支,南支由长兴岛和横沙岛分为南港和北港,南港又被九段沙分为南槽和北槽[14]。用海类型以港口航运区为主,其次为农渔业区和保留区。长江河口及海域生态保护红线面积2397.25 km2,自然保护地包括崇明东滩鸟类国家级自然保护区、九段沙湿地国家级自然保护区、长江口中华鲟自然保护区,以及陈行、青草沙、东风西沙饮用水源保护区(图1)。![]() 图 1 长江口及其毗邻海域开发利用现状Figure 1. Status of exploitation and utilization of the Yangtze River Estuary and its adjacent waters
图 1 长江口及其毗邻海域开发利用现状Figure 1. Status of exploitation and utilization of the Yangtze River Estuary and its adjacent waters1.2 数据来源及预处理
海域空间数据源自《上海市海洋功能区划(2012—2020)》和上海市已发布的生态保护红线。利用ArcGIS10.2 将上海市海洋功能区划(2012—2020)矢量数据进行栅格化处理,形成农渔业区、港口航运区、工业与城镇建设区、矿产与能源区、旅游娱乐区、河口海洋保护区、特殊利用区、保留区八类用海类型。上海市生态保护红线图经几何校正后,进行矢量化。用于构建阻力面的水深、温度、盐度、溶解氧、悬浮物浓度数据来自He等的研究[15]。
本研究用于构建阻力面的环境因子标准化方法如下:
正向因子标准化计算公式: (X−最小值)/(最大值−最小值)
负向因子标准化计算公式:(最大值−X)/(最大值−最小值)
区间因子标准化计算公式: a+[(b−a)×(X−最小值)]/(最大值−最小值)
X指各因子的实测值,最大值、最小值是指实测数据中最大值和最小值。a, b分表代表数据在规定区间的区间值。
2. 研究方法
2.1 基于MSPA的生态源地选择
形态学空间格局分析方法(MSPA)由 Vogt等基于形态学原理提出[16]。该方法是利用腐蚀、扩张、开运算、闭运算对图形进行分割、识别和分类的一种新的图像处理技术,可以识别目标像元与结构要素之间的空间拓扑关系。该方法基于土地利用数据,将土地利用类型中的具备生态功能的地类作为前景(foreground),并将其他地类作为背景(background),通过一系列的图像处理方法将前景按形态分为核心区、孤岛、孔隙、边缘、连接桥、环道和支线7种生态景观类型,使生态源地以及生态廊道的识别更科学[17-18]。本研究以农渔业区(去除农业围垦区)、河口海洋保护区、保留区作为前景数据,以港口航运区、工业与城镇建设区、矿产与能源区、旅游娱乐区、特殊利用区为背景数据,获得前景与背景的二元分析底图。在GuidosToolbox3.2平台下,二元栅格图像,采用八邻域图像细化分析法进行形态学空间分析,将研究区重新解译为核心区、环道区、孔隙、岛状斑块、边缘区、支线、桥接区七类景观[8,20]。利用Conefor 2.6软件计算景观连通性3类重要指标:整体连通性指数(integral index of connectivity, IIC) 、可能连通性(probability index of connectivity, dPC) 以及相对重要性指数(relative importance index, dI) 作为生态源地选择的重要参考[5]。选取核心区中斑块面积最大的15个源地进行连通性分析,将斑块连通距离阈值分别设置为500 、
5000 、10000 、50000 、80000 、100000 m,并据此进行阈值筛选,随着斑块连通距离的变化,IIC和dPC保持稳定基本不变时的阈值作为最终分析的距离阈值。拟合结果发现80000 m效果最好。为了与IIC结果具有可比性,将可能性概率设置为0.5,选择dPC值大于1的斑块作为重要生态源地。2.2 空间阻力面构建
春季是长江口主要鱼类的洄游季节。以长江口5月份的水深、温度、盐度、溶解氧、悬浮物浓度为基础数据,将基础数据依据本研究标准化方法进行标准化后进行克里金插值,将不同环境因子转为空间栅格数据,利用自然断点法进行重分类,并等权重叠加各因子栅格数据构建阻力面(表1)。
表 1 阻力面构建因子Table 1. Resistance surface constructors阻力因子 阻力因子属性 水深 负向因子,水深越深,阻力越低 水体温度 负向因子,温度越高,阻力越低 水体盐度 盐度在区间(5~12)时阻力最小,
越小和越大阻力值均增加悬浮物浓度 正向因子,悬浮物浓度越高,阻力越高 溶解氧 负向因子,溶解氧越高,阻力越小 2.3 廊道提取
利用Linkage Mapper(LM)工具调用软件Circuitscape,运用电路理论提取生态廊道、“生态夹点”和“生态障碍点”。Circuitscape程序基于电路理论对不同的景观进行连接建模,该模型利用电荷的随机游动特性,模拟物种个体或基因流在某一景观中迁移扩散过程[19-20]。物种个体或基因流被视为电子,景观面被视为电导面,由此模拟电导面中的全向电流,并得出斑块间的全向电流密度值,表征物种扩散概率。在整个景观中计算出的有效电阻、电流与生态过程相关联[21-22]。
①提取生态廊道。利用LM工具Linkage Pathway Tools调用Circuitscape软件将生态源地和综合阻力面导入 ArcGIS中,识别最小成本路径(least-cost path method,LCP),获得最小成本距离廊道。
②识别生态节点。生态“夹点”(又称瓶颈),是对研究区连接性具有重要作用的区域,通过LM 中Pinchpoint Mapper工具提取。生态“障碍点”是影响通道质量的重要屏障(障碍物),通过LM中 Barrier Mapper 工具提取。
3. 结果分析
3.1 基于MSPA 与景观连通性的生态源地分析
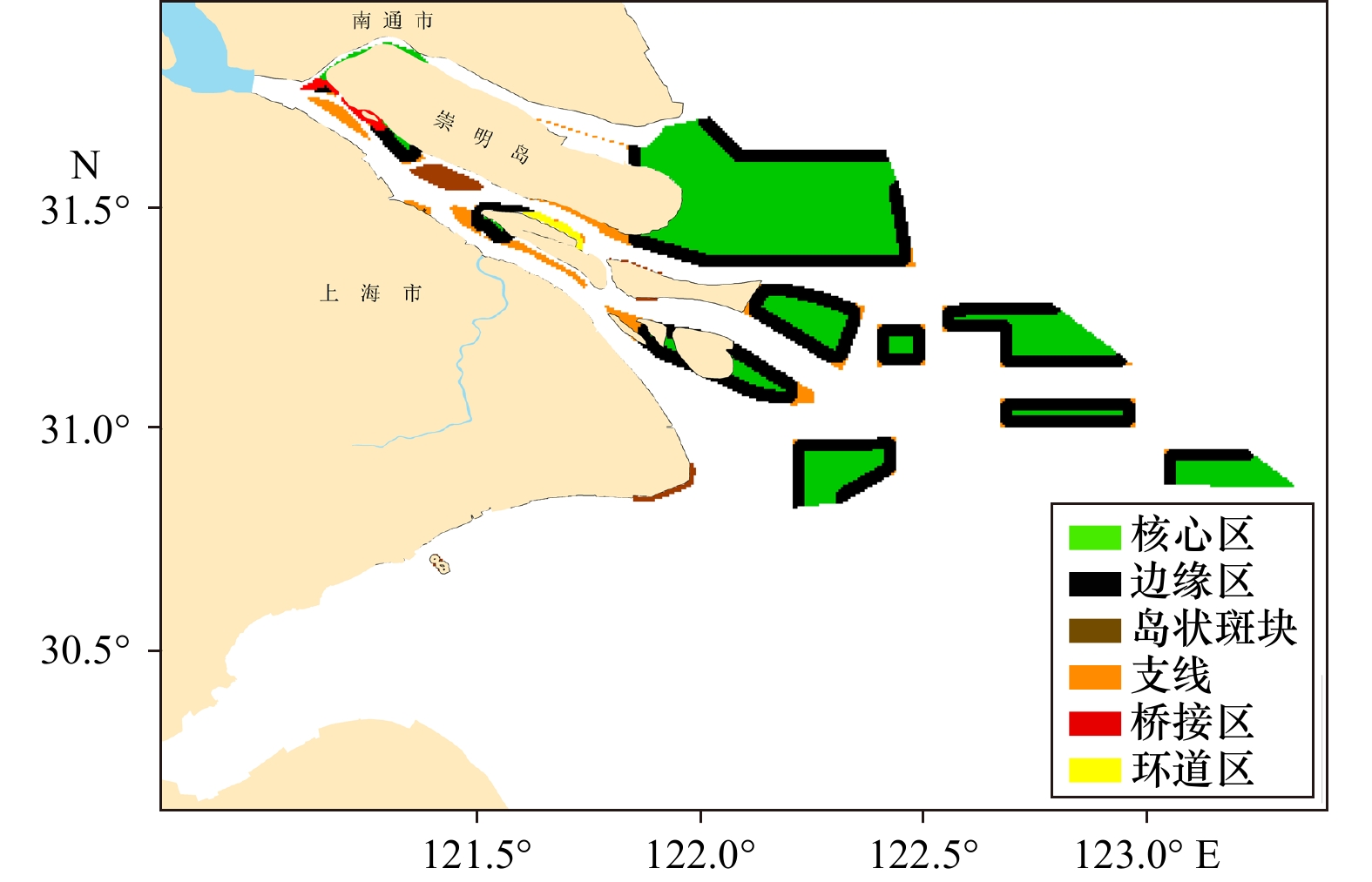
结果表明(表2、图2),长江口海域对生态保护具有重要作用的核心区景观面积约为
2050 m2,是目前长江口纳入保护区面积的3倍。边缘区是核心区的外部边缘,其面积仅次于核心区,景观斑块具有较好的边缘效应。环线和桥接区面积较小,斑块间和斑块内部的连通性较低,支线占景观总面积的6%,表明核心区与外部景观具有一定的连通作用。孤岛分布零散,所占比例仅2%,可设置为踏脚石斑块,增加生态网络稳定性。表 2 基于MSPA的景观类型统计Table 2. Statistical results of landscape types based on MSPA类型 面积/km2 占前景海洋
保护区比例/ %占前景农渔
业区比例/ %占前景保留
区比例/ %占景观要素
比例/ %核心区 2050.23 284 123 120 51 桥接区 41.07 2 2 2 1 环道区 26.75 2 2 2 1 支线 223.07 13 13 13 6 孤岛 93.56 6 6 5 2 边缘区 1611.16 97 97 95 40 ![]() 图 2 MSPA分析的长江口及其毗邻海域景观类型Figure 2. Landscape type of Yangtze River Estuary and its adjacent waters based on MSPA analysis
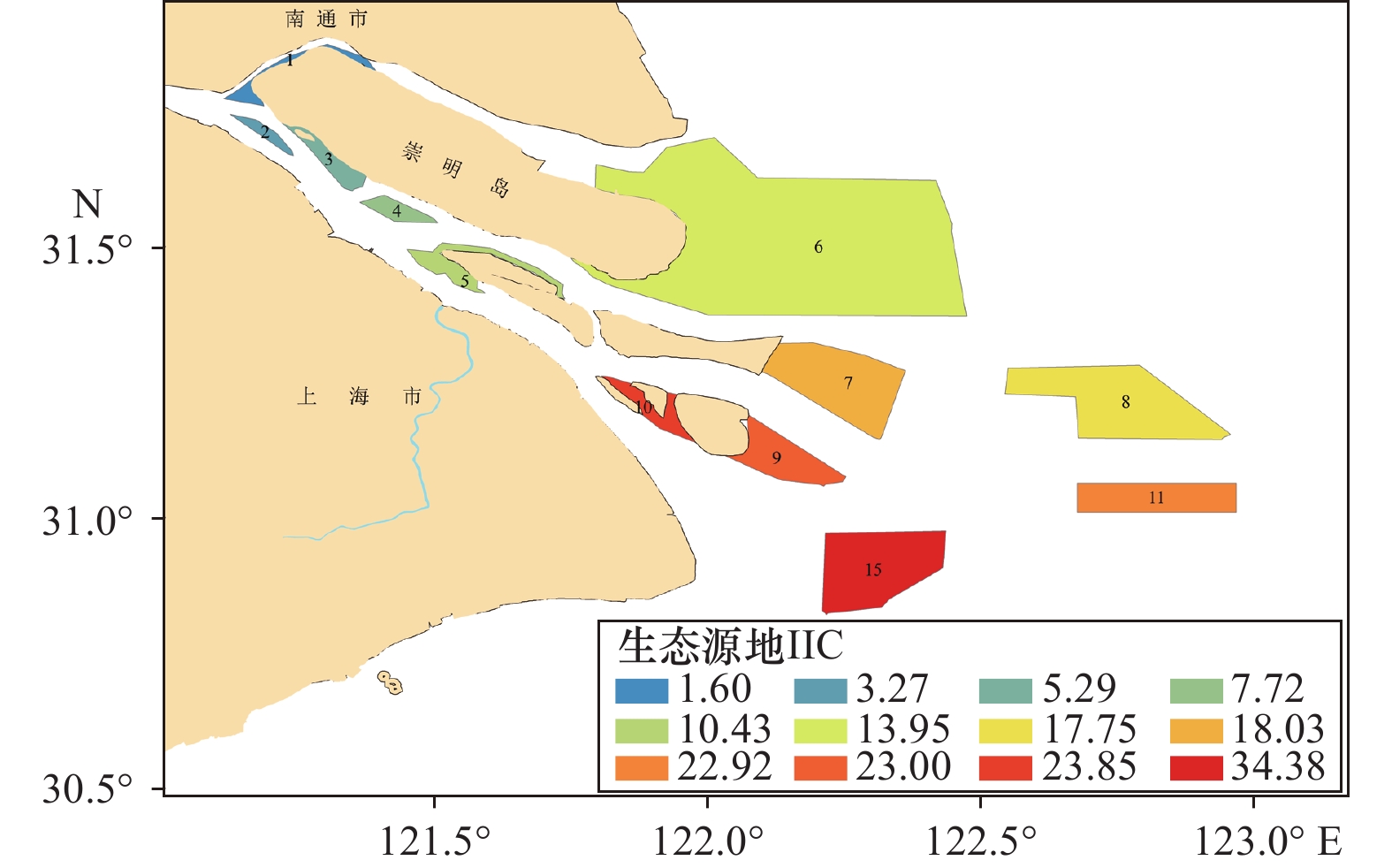
图 2 MSPA分析的长江口及其毗邻海域景观类型Figure 2. Landscape type of Yangtze River Estuary and its adjacent waters based on MSPA analysis基于dPC值筛选结果(表3、图3),长江口共筛选重要生态源地12个,总面积
3080 km2,占上海市海洋功能区划面积28.6%,占前景农渔业区、海洋保护区和保留区面积的68.0%。通过对IIC、dPC、dI指数分析,长江口九段沙及周边海域连通性指数最高,是连接长江口与杭州湾、长江口内外的重要生态源地。其次连通性重要的生态源地为横沙浅滩海域和口门外农渔业区海域。长江口内生态源地相对重要性较口门外低。但其发挥着重要桥接区的作用。表 3 生态源地重要性指数排序Table 3. Ranking of the importance index of ecological sources序号 源地编号 面积/km2 IIC dPC dI/% 1 15 284.43 34.51 34.38 33 2 11 166.97 23.13 22.92 24 3 10 56.33 22.68 23.84 23 4 9 124.25 21.30 23.00 22 5 8 359.54 17.47 17.76 19 6 7 273.67 16.57 18.03 18 7 6 1581.90 13.94 13.95 17 8 5 72.90 10.96 10.43 15 9 4 35.16 7.62 7.72 10 10 3 55.30 5.41 5.29 8 11 2 20.15 3.25 3.27 5 12 1 49.34 1.63 1.60 2 3.2 生态廊道的提取分析
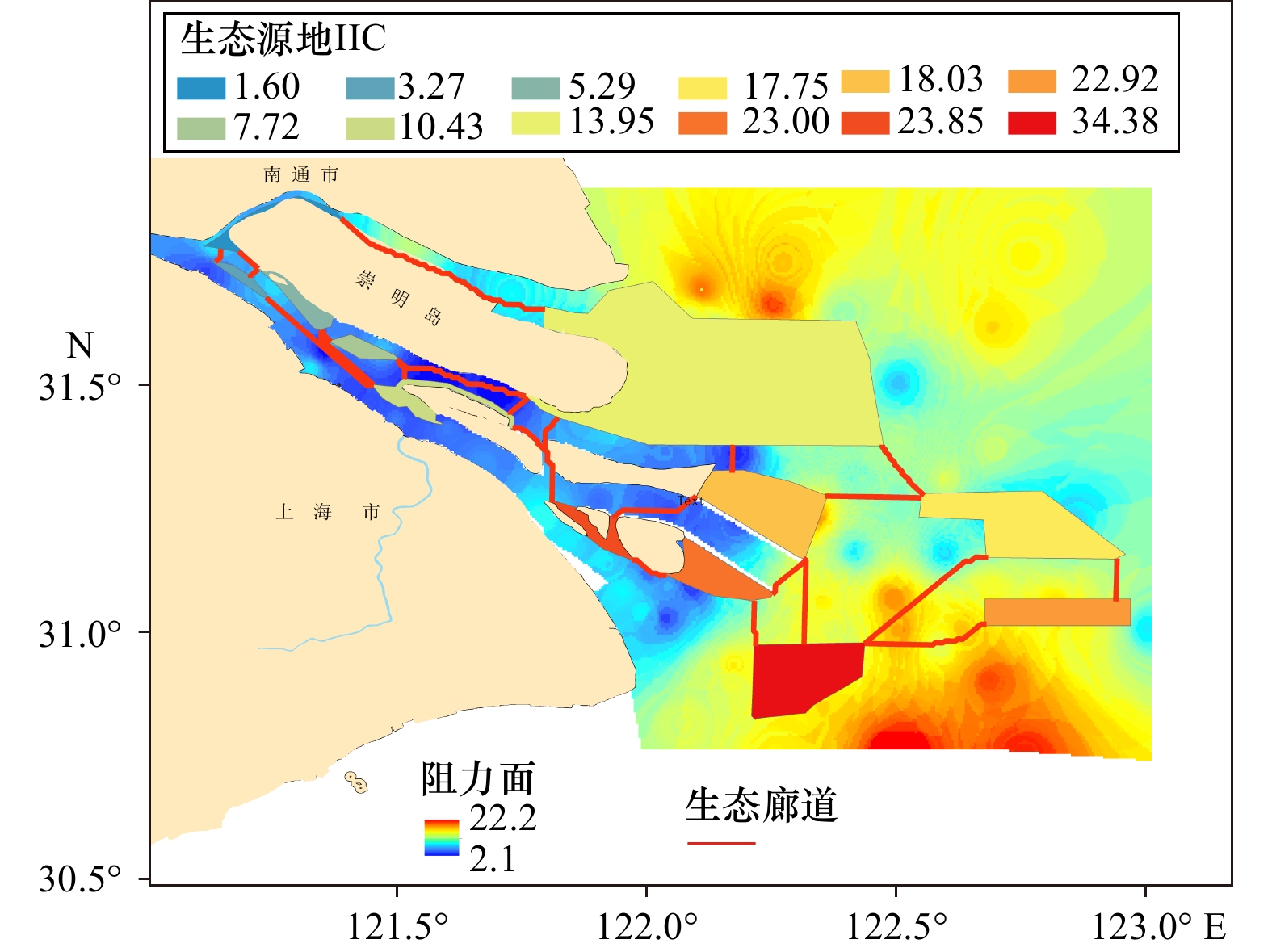
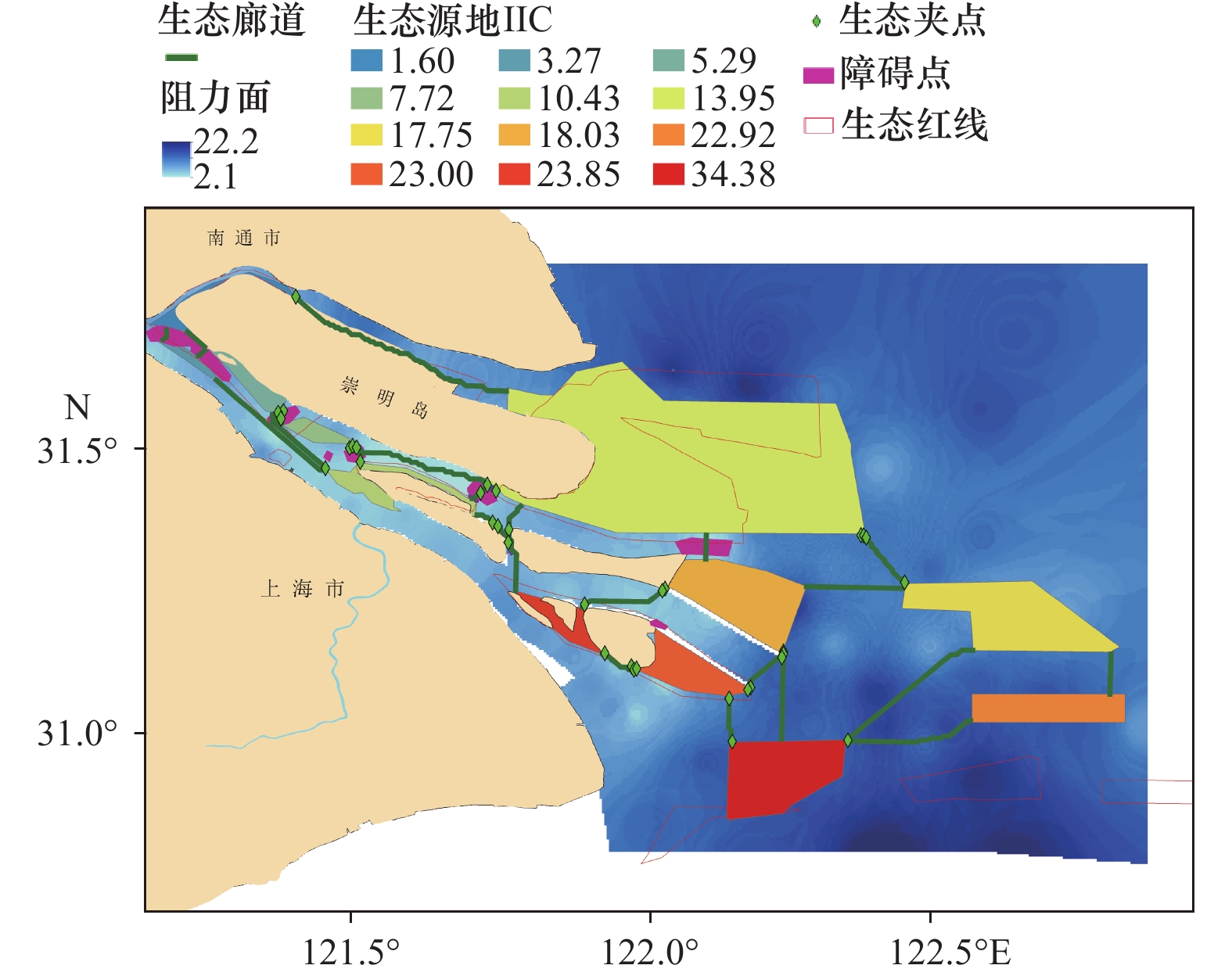
提取出长江口生态廊道23条(表4),总体上廊道主要分布于长江口南支及口门海域。其中最长廊道约47 km,为崇明北支廊道,连接生态源地1和生态源地6。阻力最高的廊道为生态源地2与生态源地5之间的连接廊道。九段沙与长兴岛、横沙岛之间的廊道阻力值也较大,不利于物种之间的扩散。生态源地15是长江口与杭州湾以及口门内外连接生物扩散的重要源地,但与其他源地间廊道阻力较高,表明物种在这些区域扩散存在一定的抑制(图4)。
表 4 生态廊道属性表Table 4. Attribute of ecological corridor序号 廊道长度/ m 廊道有效阻力/(Pa·m−1) 1 3299 2 003 2 2533 2073 3 28780 67996 4 3216 5275 5 4948 7072 6 27843 25577 7 5466 19015 8 47341 42735 9 16281 13471 10 20761 46311 11 8165 12101 12 17945 38745 13 20444 52361 14 5266 4549 15 11015 39451 16 10346 22618 17 25192 31866 18 9663 5366 19 19526 64396 20 18359 31450 21 6281 1 971 22 31396 42282 23 14514 18322 3.3 生态夹点提取分析
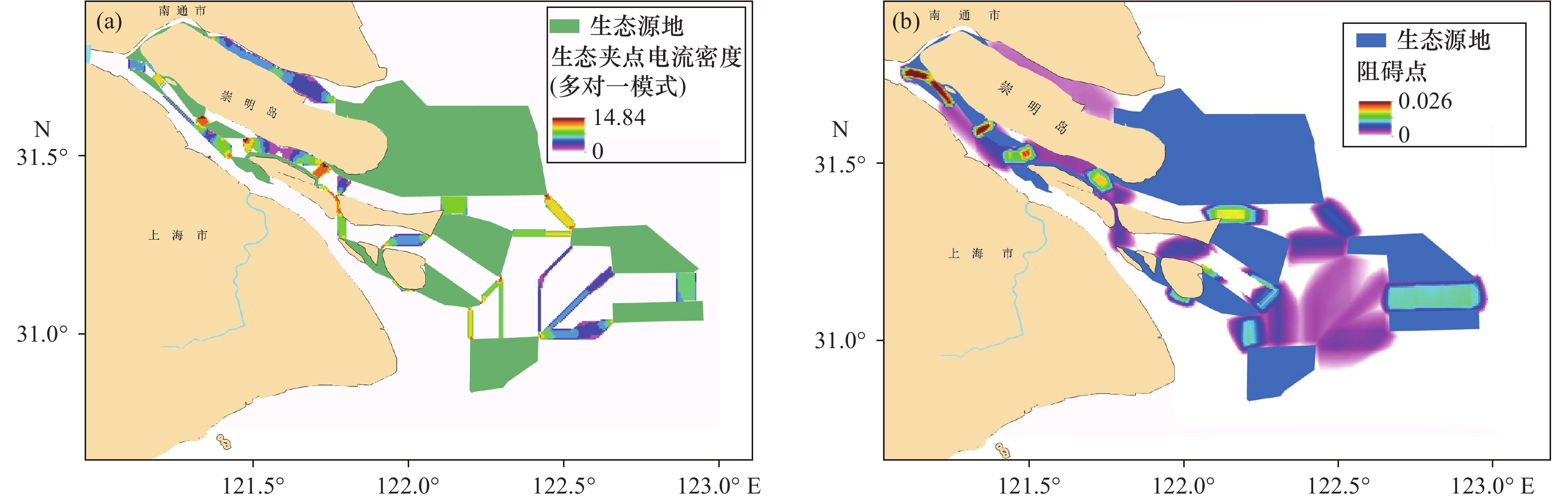
通过对生态夹点提取分析[图5(a)],长江口生态源地间的生态夹点主要分布于各斑块的廊道连接处,在长兴岛与崇明岛之间的生态夹点分布较为密集。生态障碍点[图5(b)]的提取分析表明,长江口各源地间的自然障碍点较少,连接较为通畅,生态障碍点主要分布于长江口南支崇明岛西缘。生态源地1至3之间生态障碍点较多。
![]() 图 5 长江口及其毗邻海域生态节点提取图Figure 5. Extraction map of ecological nodes in the Yangtze River Estuary and its adjacent sea areas
图 5 长江口及其毗邻海域生态节点提取图Figure 5. Extraction map of ecological nodes in the Yangtze River Estuary and its adjacent sea areas3.4 生态保护空间匹配分析
从上海市海洋生态保护红线与长江口生态源地的叠加分析可知(图6),长江口海域保护空间主要集中在崇明岛、九段沙周缘,以及口门外部分区域。海域生态红线占生态源地的面积约为80%。横沙东滩及以南支口门外毗邻重要源地15未被纳入生态红线。口门外红线区与生态源地空间匹配度不高。
![]() 图 6 长江口生态保护空间与生态源地匹配分析图Figure 6. Spatial matching of ecological protection area and ecological sources in the Yangtze River Estuary
图 6 长江口生态保护空间与生态源地匹配分析图Figure 6. Spatial matching of ecological protection area and ecological sources in the Yangtze River Estuary4. 讨论
4.1 长江口生态保护网络格局及连通性
本研究基于形态学空间格局分析(MSPA)方法,从像元层面评价长江口生态空间及其功能连通性。从源地的重要性来看,长江口南支口门附近生态源地的重要性最高。该区域处于长江口与杭州湾以及口门外毗邻海域的交汇区,对连接长江口生态系统非常重要。对长江口洄游种凤鲚(Coilia mystus)、淡水种银飘鱼(Pseudolaubuca sinensis)、海水种日本鳀(Engraulis japonicus)和河口种子陵吻虾虎鱼(Rhinogobius giurinus)鱼类的研究发现,长江口凤鲚高密度分布区主要位于北支和南支口门附近,银飘鱼最适栖息生境主要位于长江口南支以及崇明岛与长兴岛之间海域,日本鳀仔稚鱼春季和子陵吻虾虎鱼主要分布于南支口门及外毗邻海域[23-24]。盐度是影响长江口四种典型鱼类仔稚鱼的重要环境因子[15, 23]。由于长江口盐度不同月份间(5月和8月)变化较大,特别是8月丰水期河口等盐线呈现明显向东南方向凸出的特征[25]。由此可见,长江口典型洄游型鱼类凤鲚的产卵生境主要位于南支口内区域,个体在孵化后进入长江口北支、舟山群岛水域等盐度相对较低的西侧水域生活。其主要洄游路径为近岸海域成鱼进入长江口南支、北支产卵,孵化后在口门附近育幼并再进入近岸海域。此外,由于长江口南支形成冲淡水通道,盐度较低,是长江口淡水鱼类(如银飘鱼)和河口型鱼类(如子陵吻虾虎鱼)的主要生境。对长江口生物完整性指数(IBI)的研究发现,长江口南支口门与北支口门附近完整性普遍较高[26]。
对长江口凤鲚廊道的研究发现,口门区域和感潮河段是河口短距离洄游的重要生境,河口区生境多样性是由盐度等关键因子驱动,而理化性质的变化首先受河口形态(宽度、形状等)的影响[14]。这些研究均印证了长江口南支口门附近生境的重要性,这些区域主要以九段沙和本研究中的生态源地15海域组成。崇明东滩与北支附近生态源地是仅次于长江口南支源地的区域,长江口内源地1至源地3相对重要性较低,但作为河海洄游物种桥接区,起垫脚石作用。长江口生态夹点主要分布于长兴岛与崇明岛之间,这些区域对口门内物种的连通起到重要作用,该区域生境修复的相关研究表明,春夏秋三季在该区域修复区的物种数均多于对照区,主要优势物种为凤鲚(Coilia mystus)、刀鲚(Coilia nasus)、中国花鲈(Coilia nasus)、拉氏狼牙虾虎鱼(Odontamblyopus)等,鱼类食源相较于对照区更为广泛,表明物种在生境间的联系也较为广泛[27]。长江口障碍点主要分布于崇明岛西缘区域,对该区域生态源地间的连接造成较大的障碍。可能的原因是该区域沙体变化较大,不利于生境稳定。对长江口2002—2019年滩涂演变分析表明,崇明西侧上扁担沙处于自然演变状态,因无工程守护局部演变剧烈[28]。杨立君等利用遥感影像对崇明岛滩涂冲淤演变特征分析表明,10~20年后北支有可能断流,崇西沙体将可能存在较大的变化[29]。这些研究也印证了该区域生境多变。
4.2 长江口海域多尺度生态网络优化
从国际研究趋势来看,结合连通性分析识别生态廊道,构建生态网络是有效减缓甚至遏制区域生物多样性丧失和自然生态系统退化的重要途径[30]。由于许多生物不同生活史阶段需利用不同类型的栖息地[9],河口海域连通性是重要的决定因素。例如,Pelage等对巴西河口洄游鱼类银鲈(Eucinostomus argenteus)的连通性研究发现,连通性可能会影响生态系统的功能,因为它们会影响相邻栖息地之间的营养途径和能量流动[11]。Turk-Boyer等利用海洋生物耦合模型对墨西哥湾3种海洋生物(蓝蟹Callinectes bellicosus,长棘赤鳞鳃Spondylus limbatus,豹纹喙鲈Mycteroperca rosacea)的幼体扩散和种群遗传学研究揭示海洋和沿海栖息地之间的连通性,并提出生物廊道作为一个独特的管理单位对于渔业保护尤其重要[31]。Pendoley等利用地理空间方法识别出了澳大利亚大堡礁平背海龟(Natator depressus)生态廊道[32]。Krost等构建了近岸海域廊道的构建方法,识别优先保护区域,并评估了德国基尔海湾连通性[33]。从已有研究来看,生态廊道的研究大多基于特定物种生活史过程及其对海洋生境的需求来识别。本研究基于海域功能用途分区,重点研究了现有利用模式对海洋生境的重塑作用,相对于单一物种生态廊道及其连通性的评估,基于海域利用模式的评估整体上识别了重要的生态源地及其源地间最优廊道,对多物种的保护及其现有保护网络格局的优化更具指导意义。本研究提取的重要生态源地与2023年上海市公布的海洋生态保护红线具有较好的匹配性,尤其是在长江口内基本重合,表明上海市海洋管理具有较好的连续性。然而,在长江口门附近海域,本研究识别的重要源地还未纳入到上海市生态保护红线,尤其是九段沙—横沙岛东缘—崇明东滩一线相邻海域,口门内外的主要连通区域缺少保护空间,不利于长江口生态系统完整性和整体连通性。
5. 结论
长江口海域生境分布格局受到海洋功功能区划管理的深刻影响。本研究认为增加长江口九段沙—横沙岛东缘—崇明东滩一线口门海域生态空间,对促进长江口整体性保护具有重要作用,尤其是在九段沙南侧海域、横沙东滩前缘海域保护空间不足。基于MSPA的生态网络构建方法可有效用于河口水域生态系统保护网络的构建,该方法与特定物种生态廊道识别方法相结合,可支撑海洋管理分区等工作。此外,我国海域管理从海洋功能区划已转变为国土空间规划,海域“两空间一红线”(两空间为生态空间和开发利用空间,一红线为生态保护红线)划定应深入考虑海洋功能区划对生境的重塑作用。
-
![]()
图 1 长江口及其毗邻海域开发利用现状
Figure 1. Status of exploitation and utilization of the Yangtze River Estuary and its adjacent waters
![]()
图 2 MSPA分析的长江口及其毗邻海域景观类型
Figure 2. Landscape type of Yangtze River Estuary and its adjacent waters based on MSPA analysis
![]()
图 5 长江口及其毗邻海域生态节点提取图
Figure 5. Extraction map of ecological nodes in the Yangtze River Estuary and its adjacent sea areas
![]()
图 6 长江口生态保护空间与生态源地匹配分析图
Figure 6. Spatial matching of ecological protection area and ecological sources in the Yangtze River Estuary
表 1 阻力面构建因子
Table 1 Resistance surface constructors
阻力因子 阻力因子属性 水深 负向因子,水深越深,阻力越低 水体温度 负向因子,温度越高,阻力越低 水体盐度 盐度在区间(5~12)时阻力最小,
越小和越大阻力值均增加悬浮物浓度 正向因子,悬浮物浓度越高,阻力越高 溶解氧 负向因子,溶解氧越高,阻力越小  下载: 导出CSV
下载: 导出CSV
表 2 基于MSPA的景观类型统计
Table 2 Statistical results of landscape types based on MSPA
类型 面积/km2 占前景海洋
保护区比例/ %占前景农渔
业区比例/ %占前景保留
区比例/ %占景观要素
比例/ %核心区 2050.23 284 123 120 51 桥接区 41.07 2 2 2 1 环道区 26.75 2 2 2 1 支线 223.07 13 13 13 6 孤岛 93.56 6 6 5 2 边缘区 1611.16 97 97 95 40
下载: 导出CSV
表 3 生态源地重要性指数排序
Table 3 Ranking of the importance index of ecological sources
序号 源地编号 面积/km2 IIC dPC dI/% 1 15 284.43 34.51 34.38 33 2 11 166.97 23.13 22.92 24 3 10 56.33 22.68 23.84 23 4 9 124.25 21.30 23.00 22 5 8 359.54 17.47 17.76 19 6 7 273.67 16.57 18.03 18 7 6 1581.90 13.94 13.95 17 8 5 72.90 10.96 10.43 15 9 4 35.16 7.62 7.72 10 10 3 55.30 5.41 5.29 8 11 2 20.15 3.25 3.27 5 12 1 49.34 1.63 1.60 2
下载: 导出CSV
表 4 生态廊道属性表
Table 4 Attribute of ecological corridor
序号 廊道长度/ m 廊道有效阻力/(Pa·m−1) 1 3299 2 003 2 2533 2073 3 28780 67996 4 3216 5275 5 4948 7072 6 27843 25577 7 5466 19015 8 47341 42735 9 16281 13471 10 20761 46311 11 8165 12101 12 17945 38745 13 20444 52361 14 5266 4549 15 11015 39451 16 10346 22618 17 25192 31866 18 9663 5366 19 19526 64396 20 18359 31450 21 6281 1 971 22 31396 42282 23 14514 18322
下载: 导出CSV
-
[1] 肖笃宁,陈文波,郭福良. 论生态安全的基本概念和研究内容[J]. 应用生态学报,2002,13(3):354-358. XIAO D N,CHEN W B,GUO F L. On the basic concepts and contents of ecological security[J]. Chinese Journal of Applied Ecology,2002,13(3):354-358.
[2] CUNHA N S,MAGALHÃES M R. Methodology for mapping the nation’s ecological network to mainland Portugal:a planning tool towards a green infrastructure[J]. Ecological Indicators,2019,104:802-818. DOI: 10.1016/j.ecolind.2019.04.050
[3] 代继平,朱坤,周天宇,等. 基于MSPA和MCR模型的曲靖中心城市生态网络优化研究[J]. 西北林学院学报,2022,37(6):250-257. DAI J P,ZHU K,ZHOU T Y,et al. Optimization study of Qujing central city ecological network based on MSPA and MCR models[J]. Journal of Northwest Forestry University,2022,37(6):250-257.
[4] 杜箫宇,吕飞南,王春雨,等. 基于MSPA-Conefor-MCR的县域尺度生态网络构建:以延庆区为例[J]. 应用生态学报,2023,34(4):1073-1082. DU X Y,LV F N,WANG CY,et al. Construction of ecological network based on MSPA-Conefer-MCR at the country scale:a case study in Yanqing district,Beijing,China[J]. Chinese Journal of Applied Ecology,2023,34(4):1073-1082.
[5] 艾婧文,余坤勇,黄茹鲜,等. 风电项目对潜在生态廊道的影响:基于MSPA-MCR模型[J]. 生态学报,2023,43(9):3665-3676. AI J W,YU K Y,HUANG R X,et al. The impact of wind power projects on potential ecological corridors based on the MSPA-MCR Model[J]. Acta Ecologica Sinica,2023,43(9):3665-3676.
[6] 刘婷,欧阳帅,勾蒙蒙,等. 基于MSPA模型的新型城市热景观连通性分析[J]. 生态学报,2023,43(2):615-624. LIU T,OUYANG S,GOU M M,et al. analysis connectivity of urban heat island in a new-type urbanization based on MSPA model[J]. Acta Ecologica Sinica,2023,43(2):615-624.
[7] 胡西武,贾天朝. 基于生态敏感性与景观连通性的三江源国家公园生态安全格局构建与优化[J]. 长江流域资源与环境,2023,32(8):1725-1733. HU X W,JIA C T. Construction and optimization of ecological security pattern in Three-River-Source National Park based on ecological sensitivity and landscape connectivity[J]. Resources and Environment in the Yangtze Basin,2023,32(8):1725-1733.
[8] 王崑,马春旭,郑伊含,等. 基于MSPA 模型和生态保护重要性评价的市域生态空间网络构建及优化[J]. 水土保持通报,2023,43(4):220-228. WANG K,MA C X,ZHENG Y H,et al. Construction and optimization of urban ecological space network based on MSPA model and ecological protection importance evaluation[J]. Bulletin of Soil and Water Conservation,2023,43(4):220-228.
[9] SHEAVES M. Consequences of ecological connectivity:the coastal ecosystem mosaic[J]. Marine Ecology Progress Series,2009,391:107-115. DOI: 10.3354/meps08121
[10] ELLIOTT M,WHITFIELD A K. Challenging paradigms in estuarine ecology and management[J]. Estuarine,Coastal and Shelf Science,2011,94(4):306-314. DOI: 10.1016/j.ecss.2011.06.016
[11] PELAGE L,GONZALEZ J G,LE LOC’H F,et al. An Importance of estuary morphology for ecological connectivity with their adjacent coast:a case study in Brazilian tropical estuaries[J]. Estuarine,Coastal and Shelf Science,2021,251(1/2):107184.
[12] FRIESEN S K,RUBIDGE E,MARTONE R,et al. Effects of changing ocean temperatures on ecological connectivity among marine protected areas in northern British Columbia[J]. Ocean and Coastal Management,2021,211:105776. DOI: 10.1016/j.ocecoaman.2021.105776
[13] WOLANSKI E. Bounded and unbounded boundaries–Untangling mechanisms for estuarine-marine ecological connectivity:scales of m to10,000 km-a review[J]. Estuarine,Coastal and Shelf Science,2017,198:378-392. DOI: 10.1016/j.ecss.2016.06.022
[14] 陈吉余,陈沈良. 近20年来长江口生态环境变化[J]. 净水技术,2002,21(S1):1-3. CHEN J Y,CHEN S Ecological environment changes in the Changjiang River Estuary over recent 20 years[J]. Water Purification Technology,2002,21(S1):1-3.
[15] HE Y L,ZHAO L X,LIU S H,et al. Delineation of estuarine ecological corridors using the MaxEnt model to protect marine fishery biodiversity[J]. Frontiers in Marine Science,2022:DOI 10.3389/fmars.2022.966621.
[16] VOGT P,FERRARI J R,LOOKINGBILL T R,et al. Mapping functional connectivity[J]. Ecological Indicators,2009,9(1):64-71. DOI: 10.1016/j.ecolind.2008.01.011
[17] 谢名睿,危小建,赵莉,等. 南昌市生态用地景观结构与网络连通性多情景模拟[J]. 水土保持通报,2023,43(2):202-211. XIE M R,WEI X J,ZHAO L,et al. Multi scenario simulation of landscape structure and network connectivity of ecological land in Nanchang City[J]. Bulletin of Soil and Water Conservation,2023,43(2):202-211.
[18] 张启舜,李飞雪,王帝文,等. 基于生态网络的江苏省生态空间连通性变化研究[J]. 生态学报,2021,41(8):3007-3020. ZHANG Q S,LI F X,WANG D W,et al. Analysis on changes of ecological spatial connectivity in Jiangsu Province based on ecological network[J]. Acta Ecologica Sinica,2021,41(8):3007-3020.
[19] MCRAE B H. Isolation by resistance[J]. Evolution, 2006,60(8):1551-1556.
[20] DOLYE P G,SNELL J L. Random walks and electric networks[J]. arXiv,2000:DOI: 10.2307/2322439.
[21] MCRAE B H,DICKSON B G,KEITT T H,et al. Using circuit theory to model connectivity in ecology,evolution,and conservation[J]. Ecology,2008,89(10):2712-2724. DOI: 10.1890/07-1861.1
[22] MCRAE B H,SHAH V B,MOHAPATRA T K. Circuitscape 4 user guide. The Nature Conservancy [EB/OL]. [2024-01-09]. https://docs.circuitscape.org/Circuitscape.jl/latest/usage/.
[23] 林军. 基于物种分布模型的长江口仔稚鱼优势种生境特征研究[D]. 上海:上海海洋大学,2022. LIN J. Habitat characteristics of dominant species of fish larvae in the Yangtze estuary based on species distribution model[D]. Shanghai:Shanghai Ocean University,2022.
[24] 姜涛. 基于耳石形态和微化学特征的我国鲚属鱼类洄游生态学研究[D]. 南京:南京农业大学,2014. JIANG T. Study on migration ecology of Coilia in China based on otolith morphology and micro-chemical characteristics[D]. Nanjing:Nanjing Agricultural University,2014.
[25] 徐兆礼,沈新强. 长江口水域浮游动物生物量及其年间变化[J]. 长江流域资源与环境,2005,14(3):282-286. XU Z L,SHEN X Q. Zooplankton biomass and its variation in water near Changjiang Estuary[J]. Resources and Environment in the Yangtze Basin,2005,14(3):282-286.
[26] 谢志伟. 应用生物完整性指数(IBI)评价长江口生态健康状况[D]. 上海:上海海洋大学,2022. XIE Z W. Evaluation of the environmental health of the Yangtze Estuary by Index of Biological Integrity (IBI) [D]. Shanghai:Shanghai Ocean University,2022.
[27] 谢雯. 长江口渔业生境修复对鱼类群落及其基础食源的影响[D]. 天津:天津农学院,2023. XIE W. Effects of fisheries habitat restoration on fish community structure and basic food source in the Yangtze River Estuary[D]. Tianjin:Tianjin Agricultural University,2023.
[28] 张东来,宋磊. 长江口2002—2019 年滩涂演变分析[J]. 水运工程,2023(6):137-147. ZHANG D L,SONG L. Tidal flat evolution in Yangtze River Estuary from 2002 to 2019[J]. Port & Waterway Engineering,2023(6):137-147.
[29] 杨立君,李梦博,吴彤馨,等. Landsat影像的崇明岛滩涂冲淤演变特征分析[J]. 测绘科学,2023,48(5):162-172. YANG L J,LI M B,WU T X,et al. Analysis of the evolution characteristics of Chongming Island tidal flats in Landsat images[J]. Science of Surveying and Mapping,2023,48(5):162-172.
[30] 黄俊达,黄金玲,陈超劲. 以自然保护地为主体的广州市域生态网络构建[J]. 应用生态学报,2024,1:249-256. HUANG J D,HUANG J L,CHEN C J. Construction of ecological network in Guangzhou City,China with protected areas as the main region[J]. Chinese Journal of Applied Ecology,2024,1:249-256.
[31] TURK-BOYER P J,MORZARIA-LUNA H N,MARTINEZ-TOVAR I,et al. Ecosystem-based fisheries management of a biological corridor along the Northern Sonora Coastline (NE Gulf of California)[M]. AMEZCUA F,BELLGRAPH B. Fisheries management of Mexican and Central American Estuaries. Dordrecht:Springer Netherlands,2014:125-154.
[32] PENDOLEY K L,SCHOFIELD G,WHITTOCK P A,et al. Protected species use of a coastal marine migratory corridor connecting marine protected areas[J]. Marine Biology,2014,161(6):1455-1466.
[33] KROST P,GOERRES M,SANDOW V. Wildlife corridors under water:an approach to preserve marine biodiversity in heavily modified water bodies[J]. Journal of Coastal Conservation,2018,22(1):87-104. DOI: 10.1007/s11852-017-0578-5
计量
- 文章访问数: 46
- HTML全文浏览量: 14
- PDF下载量: 14
